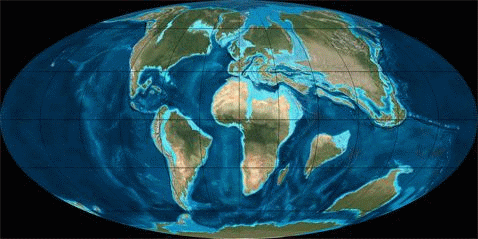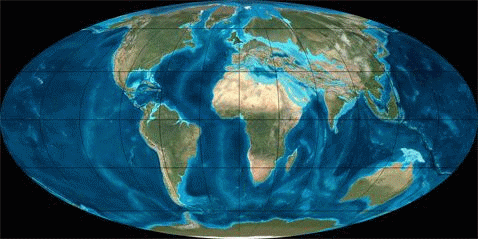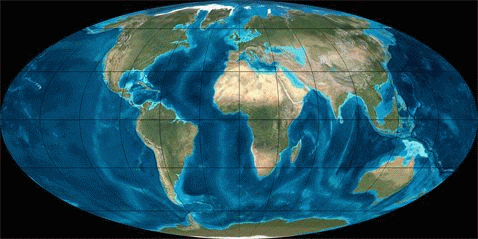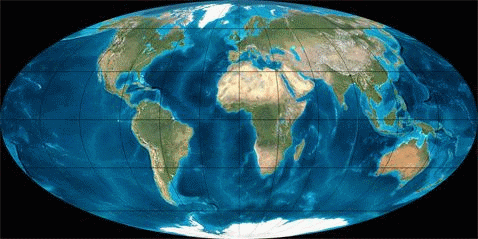
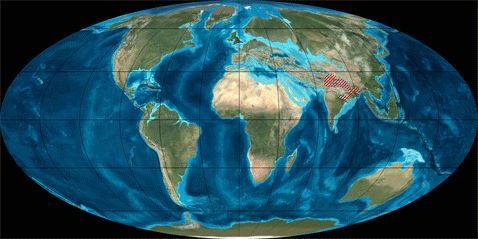
Рис. 1. Динамика литосферных плит в Кайнозое (начало 65 млн. лет назад; длительность 65 млн. лет)
Вот, например, что говорит Роберт Д. Мартин, вице-президент академического отделения в Полевом Музее и соавтор исследования, результаты которого были опубликованы в журнале Nature 18 апреля 2002 года:Применив новый метод статистического анализа данных по отношению к приматам, они обнаружили, что этот отряд млекопитающих появился не 65 миллионов лет назад, как считалось, а на 20 миллионов лет раньше. Но тогда и граница появления первых человекоподобных существ должна быть отодвинута с общепринятых 5 млн. лет до 8 млн. лет назад. Пересмотр границ происхождения, предлагаемый сотрудниками Полевого Музея, имеет ключевое значение для построения эволюционного дерева приматов, включая время появления первых людей.
«Текущие эволюционные теории развития приматов и человека зашли в тупик потому, что палеонтологи целиком положились на прямые археологические данные об ископаемых останках. <…> Наши расчёты указывают, что мы имеем ископаемые доказательства только для 5% всех исчезнувших приматов, так что работа палеонтологов по построению эволюционного дерева для отряда приматов, сравнима с тем, как если бы пришлось восстанавливать вазу, разбитую на сто осколков всего по пяти кусочкам».
«Более раннее происхождение приматов даёт объяснение ранее непонятным фактам их межвидовой раздробленности. Объяснение оказывается довольно простым, – этому способствовал дрейф континентов и последовавшее территориальное размежевание».Статистический анализ – инструмент, конечно, мощный, но ... при работе с репрезентативной выборкой из массива данных. А вот можно ли считать репрезентативной 5% выборку, да ещё и выполненную по принципу поиска под фонарём утерянного в темноте кошелька (потому, что под фонарём светло) – большой вопрос. Кто-то случайно, например, при рытье котлована под фундамент, наткнулся на древние кости или черепки (зажёг фонарь), сюда, как ночные бабочки, слетелись археологи и стали копать. Но почему здесь, а не на километр или сто километров в сторону? Да потому, что здесь светло! И вот на основе таких фактов делаются далеко (порой, очень далеко) идущие выводы.
К примеру, вынесенная в эпиграф фраза широко использовалась журналистами второй половины прошлого века в ироническом смысле, в качестве издёвки над научно-техническими достижениями СССР. Но вот недавно археологические находки показали, что мастодонты – непосредственные предки и южных слонов, и северных мамонтов, – обитали на равнинах Евразии от Карпат до Хингана. Ушедшие в тёплые края их потомки стали слонами, а ушедшие на север – мамонтами. Получается, что если и не сама Россия, которой тогда просто не было, то её территория и есть родина слонов. То есть ироническая фраза стала исторической! Да и достижения СССР задним числом оказались гораздо больше, чем это казалось досужим журналистам.Учитывая, что указанные Робертом Д. Мартином 5% тоже из серии «3p» (палец, пол, потолок), что общая площадь раскопок почти бесконечно мала по сравнению с площадью суши, на которой могли обитать наши предки, предлагается зайти «с другого конца» – попытаться логикой эволюции отыскать «перспективные» для палеонтологии места. И копать «по системе», так сказать, по методу Шлимана. Благо дарвиновское «Происхождение видов» теперь дополнено пониманием лежащих в его основе механизмов, прежде всего, мутагенеза и тектоники плит.
Иначе говоря, вообще исключить из рассмотрения ископаемые останки и заняться чисто логическими построениями. Понятно, что историческая ценность этих построений мало отличается от средневекового теологического диспута о количестве ангелов на острие иглы, но, как ни странно, она может иметь прогностическую значимость, если, конечно, в основу логических цепочек заложены верные предпосылки. А поскольку исходные посылки играют существенную, хотя и не решающую роль (из ложных посылок тоже можно получить истину), отнесёмся к их выбору со вниманием.Предпосылки. В выборе предпосылок будем исходить из дарвиновской теории происхождения видов, дополненной современным пониманием её механизмов – мутагенеза и естественного отбора. Прежде всего, отдадим себе отчёт в том, что жизнь – информационный способ наследования структуры – это естественный процесс, подверженный внешним шумовым воздействиям, начиная от теплового шума и заканчивая космическими излучениями, способными внести помехи в исходную информацию. Даже один из наиболее простых способов наследования структуры – кристаллизация – никогда не бывает бездефектным. В любом кристалле присутствуют включения не основных элементов, дислокации кристаллической решётки, наконец, в нём могут присутствовать радиоактивные элементы со своими превращениями.
Тем более это справедливо для биосинтеза – многозвенного процесса, включающего в себя многократное «чтение» генетической информации, её репликацию, «сборку» белков, их последующие конфигурационные перестройки, причём в каждом звене возможны сбои и ошибки. Поэтому фенотип всегда несколько отличается от генотипа, а, следовательно, каждый живой индивидуум несколько отличен от своих «собратьев», в том числе, и функционально. Его репродуктивные элементы также являются результатом биосинтеза и даже только поэтому воспроизводят исходную генетическую информацию не совсем точно, то есть содержат «мутации».Сразу оговорим, что мутацией будем называть любое изменение генетической структуры независимо от его локализации и внешнего проявления/непроявления в фенотипе. Присутствие так называемых «мутагенных факторов» может увеличить частоту мутаций по сравнению с некоторой фоновой, отличной от нуля даже при полном отсутствии этих факторов (так, даже при температурах вблизи абсолютного нуля возможны превращения элементов в радиационном распаде со всеми вытекающими последствиями).
Живые организмы, подвергаясь воздействию извне (со стороны среды обитания) и изнутри (со стороны постоянно возникающих мутаций), выработали многоступенчатые системы нейтрализации этих возмущений, начиная от ферментов-репараз и заканчивая естественным отбором (по принципу – если зла не избежать, то надо хотя бы извлечь из него пользу).Таким образом, биоэволюция – процесс взаимодействия трёх факторов: среды обитания, мутаций и репарационных систем живых организмов. Отсюда и её кажущаяся целенаправленность (понятно, что генные инженеры, как и создаваемые ими мутации не относятся к естественным факторам). С учётом изложенного можно сформулировать два постулата:
Постулат 1. Естественные мутации не целенаправлены.Следствия. По сути, биоэволюция представляет собой «изобретённый» задолго до появления вариационного исчисления (да и появления математиков как таковых) вариационный метод поиска оптимума. Биоэволюция точно так же нащупывает путь посредством малых отклонений, оценивает результаты, фиксирует наилучший и делает следующий шажок. Отсюда:
Постулат 2. Мутации живых организмов происходят всегда.
Следствие 1. Прогностическая функция биоэволюции тождественно равна 0. Из множества мутаций закрепляются в потомстве только те, которые наилучшим образом удовлетворяют текущим условиям обитания. Работая «от достигнутого» эволюция не может определять и учитывать, что какая-то из отвергнутых мутаций ещё через несколько шагов могла бы дать гораздо больший эффект, чем избранная. Точно так же, как и приспосабливаться к ещё не наступившим изменениям среды обитания.Обсудим эти следствия подробнее. Первое следствие исключает «тактические отступления» эволюции, то есть закрепление в потомстве изменений, ослабляющих приспособленность организма к текущим условиям в расчёте на будущие достижения. Говоря проще, эволюция следует «золотому» правилу альпинистов – «не терять высоты» – каждый эволюционный шаг приближает вид к оптимуму в текущих условиях среды обитания. Опережающие время «изобретения» эволюции «гаснут» до лучших времён. К примеру, «млекопитание» потомства двумя видами амазонских рыб (арапаима и дискус), не было применено ни другими рыбами, ни земноводными, ни пресмыкающимися и вновь было «открыто» млекопитающими.
Следствие 2. Постоянство условий обитания ведёт к диссипации видов. В самом деле, при неизменных условиях обитания оптимум рано или поздно будет достигнут, но согласно 2-му постулату мутации не прекратятся. Среди них найдутся такие, которые обеспечат примерно такой же оптимум, но в изменённом фенотипе. Возникнет подвид ... В конечном итоге биоэволюция в стабильных условиях обитания приведёт к расщеплению исходного вида, заполняя все мельчайшие уголки и щёлочки экологической ниши.
Второе следствие ещё более существенно. Его подтверждением может служить ряд хорошо известных фактов. Это и благополучно дожившее до наших дней живое «ископаемое» – целакант, демонстрирующий стабильность вида в стабильных условиях. Это и класс коралловых полипов, десятки миллионов лет строящих свои «бастионы» и за это время создавших на единой основе разнообразие видов, подвидов, семейств и т.д., практически нацело заполнив свою экологическую нишу.Важность же его для нашей темы в том, что радикальные изменения вида происходят лишь при изменении условий среды обитания. Именно в этом случае ранее достигнутый оптимум становится несостоятельным, вид ставится на грань выживания и вынужден вымереть (что и происходило многократно), если не произойдёт мутация, радикально меняющая его в нужную сторону. Таким образом, появление существенно новых типов, классов, видов и т.д. требует длительного постепенного изменения среды обитания на грани вымирания.
С этой точки зрения представляет интерес эволюция вида с учётом тектоники плит, то есть при монотонном изменении условий обитания. Возможны два направления таких изменений: монотонное смягчение условий (например, при движении материка к экватору) и прямо противоположное – ужесточение условий (материк движется в высокие широты).В первом случае вид, будучи жизнеспособным в более суровых условиях, в новых, более мягких, не нуждается в своём совершенствовании. Более того, мутации, делавшие особь мало жизнеспособной, а порой и летальные, теперь могут оказаться приемлемыми, соответствующие мутанты оказываются репродуктивными и оставляют потомство. Вид «расщепляется» на подвиды, семейства и т.п.
В случае ужесточения условий обитания всё происходит наоборот, – выживают самые приспособленные, самые эффективные в новых условиях особи. Многие подвиды (порой целые виды и семейства, утратившие возможность адаптации) вымирают. Такой вывод, например, был сделан авторами статьи в журнале Science об исследовании морских двустворчатых моллюсков. Исследователи анализировали базу данных останков морских двустворчатых моллюсков, обнаруженных к настоящему моменту. Раковины этих существ очень прочны и способны сохраняться в течение миллионов лет.Изучив в общей сложности 1678 типов двустворчатых моллюсков, обитавших на Земле в течение последних 200 миллионов лет, авторы работы выяснили, что в периоды массового вымирания, которые случались в истории планеты несколько раз, исчезали не отдельные виды, а целые группы «родственников».
В рамках рассматриваемой узкой темы мы не будем касаться вымирания моллюсков или динозавров и других биокатастроф. Читатель может сам сопоставить их с данными по тектонике плит, мы же ограничимся здесь только Кайнозоем (последние 65 мил. лет), в течение которого зарождалась и развивалась современная фауна.В таблице 1 грубыми мазками показано развитие животного мира в течение Кайнозоя, а на рисунке 1 – движение литосферных плит в этот же промежуток времени (с интервалом ~ 10 млн. лет).
|
|
(млн. лет) |
(млн. лет назад) |
|
| Современная эпоха |
|
|
Современный человек, современные животные. |
| Плейстоцен |
|
|
Первобытный человек; вымирание мастодонтов и других крупных млекопитающих. |
| Плиоцен |
|
|
Сокращение разнообразия млекопитающих. |
| Миоцен |
|
|
Максимальное разнообразие млекопитающих; возникновение современных хищных зверей. |
| Олигоцен |
|
|
Увеличение разнообразия млекопитающих современного типа. |
| Эоцен |
|
|
Вымирание ранних млекопитающих. |
| Палеоцен |
|
|
Многочисленные ранние плацентарные; птицы. |
Отметим также и то, что нижняя граница появления первых человекоподобных существ совпадает со временем сокращения разнообразия млекопитающих в плиоцене. Именно тогда Индийский треугольник, продолжая свой дрейф на север, окончательно «въехал» в Евразию, породив Гималаи, Тянь-Шань и другие горные системы на стыке плит и отгородив равнины Евразии от тропического пояса. К тому же в это время с севера началось наступление ледника. В Африке всё это время климатические условия оставались стабильными.Происхождение человека. Вот теперь настало время поговорить собственно о происхождении человека. Широкораспространённый тезис о его африканской прародине не выдерживает никакой критики, прежде всего, потому, что в течение последних тридцати миллионов лет географическое положение, а, следовательно, и климатические условия Африки были неизменны. Они способствовали диссипации видов, породили огромное разнообразие приматов (примерно то же происходило в это время в Центральной Америке), в том числе, различных питекантропов, как одно из ответвлений, но отнюдь не могли порождать что-либо радикально новое.
Рассуждения о наступлении саванны на джунгли, вынудившее наших предков к прямохождению, противоречит «принципу оптимума» – полезности каждого эволюционного шага. Переход к прямохождению с открыванием врагу самой незащищённой части – живота, оправдан только при наличии достаточно развитого мозга. В то же время развитие мозга без его немедленного применения невозможно, то есть здесь налицо то самое «тактическое отступление» в расчёте на «светлое будущее». Такие шаги в эволюции принципиально запретны.А вот для того, чтобы дальнейшие поиски нашего пращура стали успешными, необходимо вести их на таком пути эволюции приматов, на котором каждый эволюционный шаг давал бы преимущество в борьбе за выживание. По нашему мнению, именно такой путь, встречая яростное сопротивление и дарвинистов, и их антиподов, почти сорок лет пробивает идея российского учёного Л. И. Ибраева, предложившего искать предков человека среди приматов, обитавших в прибрежных палеолесах типа теперешних мангров.
В настоящее время в мангровых лесах Калимантана обитают приматы «носачи», которые значительную часть времени проводят в воде. Эти обезьяны великолепно владеют прямохождением, хотя их мозг гораздо примитивнее мозга шимпанзе, волосяной покров у них развит только на голове, зато, как и у человека, развит слой подкожной жировой клетчатки, пот содержит много соли и т.п.Предлагаемые Л.И. Ибраевым в качестве наших предков наяпитеки (пока так и не найденные Pithecus naias) обитали в неогене по берегам рек, ручьёв, озёр и других пресноводных водоёмов в лесостепной предгорной местности. Занимались ловлей и собиранием моллюсков, раков, лягушек, черепах, грызунов, птичьих яиц, прибрежных ягод, фруктов и других плодов, корений и насекомых и использовали для ловли и вскрытия раковин и панцирей расколотую гальку, палки и кости. Вода и занятость передних лап препятствовали опусканию на четвереньки. Дно мелководий, часто мягкое, требовало больших плоских ступней.
Гипотеза Ибраева полностью соответствует «принципу оптимума» – каждый эволюционный шаг давал виду немедленный выигрыш, поэтому его «наяпитеки», сходные с современными носачами, заслуживают самого пристального внимания. Легко видеть, что переход к прямохождению позволял им расширить зону добычи пищи, в то же время нападать на незащищённый живот в их среде обитания было некому. Майкл Бейджент указывает ещё на одно сходство между носачами и людьми – «опущенную гортань», – хорошо известную биологам особенность человека, отличающую его от всех сухопутных животных (и чуть ли не акт «божественного творения» по мнению клерикалов). Но именно так устроено горло не только у людей, но и у дельфинов, дюгоней, китов, то есть у водных млекопитающих (и у носачей). Короче говоря, у тех, кому требуется сделать мощный быстрый вдох перед погружением или после погружения в воду. Видимо, именно в этом эволюционный смысл «опущенной гортани».Таким образом, если принять гипотезу Ибраева, предки человека – наяпитеки – обладали физиологической возможностью богатой акустической артикуляции, способной обеспечить членораздельную речь, задолго до соответствующего развития головного мозга, и получили её «бесплатно», в качестве приложения к «болотному» образу жизни. А вот объяснить подобное устройство гортани, а следовательно, и членораздельной речи, при «саванном» происхождении весьма затруднительно.
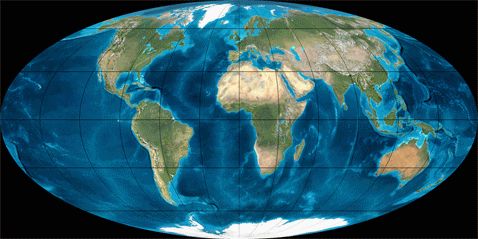
Отказываясь от «саванного» происхождения, мы вынуждены обратиться к тектонике плит, чтобы найти такое место на материках Земли, где условия обитания монотонно в течение длительного времени ужесточались, а находящиеся там животные не могли бы «эмигрировать в лучшие края». Такие условия в те времена существовали только в одном месте – к северу от места контакта Индии с Евразией после того, как их сближение начало «выращивать» хребты на месте стыка. Все остальные материки во времена зарождения наших предков в меридиональном направлении не перемещались.Более того, существовавшие раньше на этом стыке низменные, болотистые места (см. рис. 2) вполне были пригодны для реализации гипотезы Ибраева (возможно, нынешние носачи – уцелевшие в стабильных условиях на южном фланге прародины остатки наяпитеков). А вот остальные, оттесняемые растущими горами всё дальше к северу, были вынуждены приспосабливаться ко всё более жёстким условиям обитания. А тут ещё ледник с севера!
К аналогичным выводам пришли и новосибирские генетики (Л.В. Осипова и её сотрудники), которые совместно с учёными США, Бразилии и Австралии опубликовали два важных исследования по рассматриваемой здесь проблеме. Они изучили изменчивость Y-хромосомы у ряда сибирских народов, в том числе бурят, алтайцев, кетов, в сравнении с народами Северной и Южной Америки и других континентов. Оказалось, что наибольшее сходство с америндами показывают кеты – маленький народ, ныне живущий вдоль среднего течения реки Енисей, но происходящий, как предполагают антропологи, из Алтае-Саянского нагорья.Итак, можно представить себе следующую гипотетическую цепочку событий, приведшую, в конце концов, к появлению и кроманьонцев, и нас с вами:
1. Австралийская плита в своём повороте, а Африканская – в поступательном движении примерно 30 млн. лет тому назад соприкоснулись с Евразией своими континентальными шельфами.При этом Индийский треугольник и северная половина Африки оказались на экваторе. Процесс смягчения климатических условий на них достиг максимума. Для приматов наступили самые благоприятные времена, отбор практически прекратился, количество разновидностей (подвидов, видов, семейств и т.п.) начало резко расти, так как очень немногие мутации теперь делали особь нежизнеспособной или плохо приспособленной к среде обитания. Появились разнообразные питекантропы.
2. Индийский треугольник (около 20 млн. лет тому назад) соприкоснулся с Евразией континентальными частями.В мангровых лесах, выросших на мелководьях на стыке Индийского треугольника с Евразией появились наяпитеки. Условия их существования (отсутствие в манграх крупных хищников и необходимость ходить по мелководью) способствовали прямохождению, а необходимость нырять за добычей опусканию гортани.
3. Движение плит продолжается в прежнем направлении со скоростью более 5 см в год, начинается субдукция («подныривание» одной плиты под другую), которая ведёт к Альпийскому периоду горообразования.Север Африки и Индийский треугольник сдвигаются в субтропики, климатические условия на рассматриваемых территориях начинают монотонно ухудшаться, естественный отбор становится всё более жёстким, появляются, исчезают и заменяются другими различные модификации питекантропов (понятно, что аналогичным процессам подвержены не только приматы, но на других животных мы отвлекаться не будем).
Наяпитеки, поджимаемые с юга растущими горными цепями, вынуждены мигрировать в широтном направлении на запад, в сторону будущего аравийского полуострова и в Африку, и на восток, в Юго-Восточную и Восточную Азию. Те же из них, которые оказались внутри Гималайской дуги, оттеснялись этой дугой к северу, в субтропики и далее.Именно эти три направления миграции и образовали центры той «мультирегиональной эволюции человека», о которой говорит директор Института археологии и этнографии Сибирского отделения РАН академик-секретарь Отделения историко-филологических наук РАН А.П. Деревянко: «С моей точки зрения, можно выделить три модели формирования человека современного типа и его культуры. Одна реализовывалась в Африке, другая - в Юго-Восточной и Восточной Азии, третья – в Евразии (на Ближнем Востоке, Балканах, на Дону и т.д.)...»
4. Горные цепи в зоне субдукции растут и загораживают равнины Евразии от экваториального теплопритока (а субтропики и тропики от северных холодов) всё более ощутимо. Уровень суши на стыке плит повышается, сохнут болота, образуется сухопутный мост с Африкой.Мангры к югу от растущих горных цепей превращаются в джунгли, а севернее – в сухие горные леса и лесостепи. Большинство питекантропов, включая наяпитеков, превратившихся к этому времени в антропоидов (H.ergaster, H.erectus, H.georgicus и т.д.), оказавшись в джунглях (где их прямохождение не только не помогает, напротив, вредит), вымирает. Несколько легче положение «северян». Здесь двуногость и зачатки второй сигнальной системы, полученные антропоидами от наяпитеков, оказываются полезны и получают стремительное развитие.
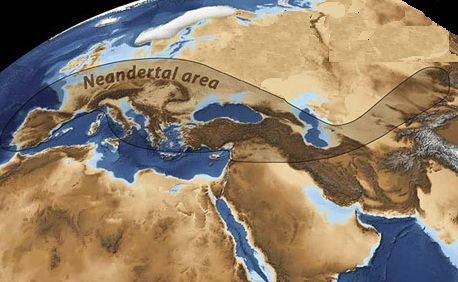
5. Следует череда оледенений, к которым «говорящие» антропоиды приспосабливаются быстрее других. Горные цепи на стыке плит надёжно прикрывают субтропики от северных холодов, поэтому там климат не меняется.Оледенения делают климат в степях Евразии всё более суровым, а отбор всё более жёстким. Процесс эволюции резко ускоряется, здесь выживают только двуногие потомки наяпитеков, «вооружённые» второй сигнальной системой. Это известный нам «неандерталец», недавно открытый «денисовец» (Homo altaiensis), а, возможно, и другие, ещё не найденные. Неандертальцы заселяют Евразию и север Африки (Рис. 4). Вторая сигнальная система позволяет обмениваться опытом и обучать молодое поколение практическим навыкам. Новые приёмы, найденные кем-то из племени, быстро становятся общим достоянием.
6. В верховьях Енисея одно из маленьких племён изобретает письменность (подробнее). Это будущие кроманьонцы, а их изобретение решает судьбу и неандертальцев, и нас с вами.Роль этого изобретения – письменности – невозможно переоценить: ведь 12 самок (а все современные люди, независимо от расы, имеют всего 12 типов митахондриальных ДНК, тех, которые передаются потомству в яйцеклетке, то есть мы все происходим от «двенадцати праматерей») это всего-навсего одна небольшая стая. И вот потомство этой маленькой стаи, обладавшее похожим на своих соседей-противников строением, примерно теми же орудиями и оружием, захватило всю землю! Безусловно, они были потомками «наяпитеков» и обладали членораздельной речью, следовательно, развитой второй сигнальной системой, но решающим фактором была именно письменность, умение хранить опыт поколений!

 |
Искать предков человека следует не в Африке, Австралии или Центральной Америке, а на юге Сибири, в северных предгорьях Алтая и среднеазиатских горных систем. Именно там потомки наяпитеков стали неандертальцами, с пользой применив и бипедализм, и опущенную, управляемую мозгом гортань, – необходимое условие членораздельной речи и второй сигнальной системы. Отсюда же шли волны переселений и в Америку, и в Европу, и Африку |