|
Здесь интересно
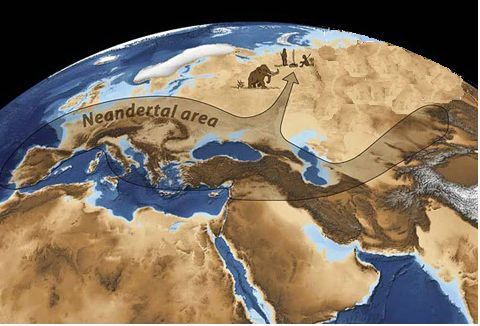
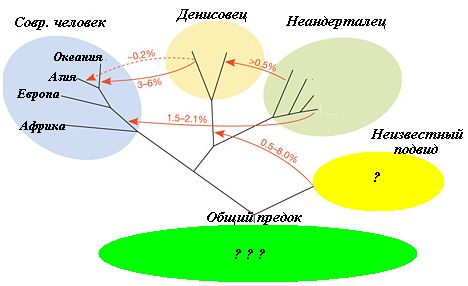
и то, что, видимо, существовал и четвёртый подвид Хомо, до сих пор не обнаруженный,
возможно, потому, что антропологи до последнего времени старательно «копали»
Африку, к которой он отношения не имеет. Понять их можно – копать интересно
там, где вероятность находок велика и где раньше коллеги уже что-либо находили.
Вот только костей больше всего бывает на кладбищах или там, где действуют
смертоносные факторы (вулканические газы, самумы и т.п.).
|