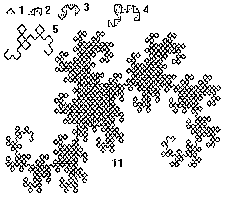
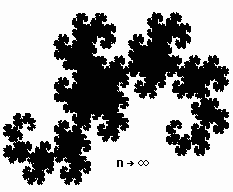
Рис. 1. Первые 5 и 11-е поколения дракона Хартера-Хейтуэя и результирующее поколение.
Но ко времени Резерфорда физика уже давно и успешно перешла в стадию обработки и анализа накопленного, а другие науки еще не нашли фундаментальных фактов, позволяющих это сделать. Так до середины прошлого века обстояло дело и с биологией. Фактов, причем не требующих даже особого труда для их добывания, было очень много, но не хватало главного – фундамента для анализа – понимания сути процессов.Положение действительно было "филателистическое". Существовала огромнейшая "коллекция", её нужно было систематизировать, но по каким принципам? И нужно отдать дань глубочайшего уважения Линнею, Ламарку, Дарвину, Менделю и другим биологам, которые зачастую ощупью нашли закономерности, в большей своей части справедливые и сейчас.
Конечно, с теперешних позиций хорошо видны их ошибки, но нельзя допустить, чтобы с одной стороны, эти ошибки заслонили достижения, а с другой, - были "канонизированы". Догматизм, особенно свойственный не точным наукам, требующий принимать на веру слова "отцов-основателей", страшнее любых ошибок.В последние годы особенно рьяной критике подвергается дарвиновская теория эволюции. На первый взгляд, в этой критике много верного, но дело в том, что эта верная критика относится не столько к Дарвину, сколько к приписываемым ему и его сторонниками, и противниками догмам. И те, и другие понимают теорию эволюции в меру своих способностей, далее свое понимание "присваивают" Дарвину и с пеной у рта защищают или опровергают. Самое примечательное здесь то, что реальные ошибки Дарвина остаются не замеченными.
Одной из таких ошибок, простительной Дарвину в его время, но непростительной сегодняшним догматикам, является мнение о непрерывности эволюции, о монотонном развитии.Увы, это не так. Весь биосинтез, и мутагенез в том числе, принципиально дискретен. Да, структура белковой молекулы однозначно определена ее первичной структурой, последовательностью аминокислот, но рабочие ее свойства зависят от более сложных уровней организации молекулы, то есть от того, как эта линейная цепочка свернулась, как соединилась с другими белковыми молекулами. И здесь уже проявляется дискретность: в первичной структуре порой можно поменять местами десятки аминокислот, но это никак не скажется на работоспособности белка, а смена одной, ключевой, полностью выведет его из строя.
Тем более это справедливо для работы механизма наследственности, что наглядно проявляется на врожденных уродствах - они, если можно так выразиться, имеют "блочный" характер, например, шестой палец, или отсутствие мягкого нёба и т.п. И всё потому, что в каком-то гене ошибочно закодирована другая аминокислота. Ведь все здоровые клетки многоклеточного организма содержат совершенно одинаковый генетический материал, а дифференциация клеток состоит в том, что в одних клетках работает одно подмножество единого генетического множества, в других - другое подмножество все того же множества. Поэтому измененный ген проявит себя везде, где он входит в активное подмножество.А вот дальше - по Дарвину: естественный отбор уничтожает неудачные мутации, а полезные закрепляет в потомстве. Бессмысленно искать "промежуточные звенья" с непрерывным, плавным увеличением мозга или укорочением передних конечностей "по миллиметру". Здесь Гулд и Элдридж правы - "недостающие звенья" не удается найти, потому что их не было. Эволюция априорно скачкообразна.
Второй существенной и опять-таки не замеченной ни сторонниками, ни противниками Дарвина ошибкой является подспудное, почти подсознательное, но пронизывающее всю теорию представление о целенаправленности эволюции.Между тем, эволюция ничуть не более целенаправленна, чем, скажем, течение реки. И как речные воды не "стремятся к морю-океану" или другому водоему, а всего лишь оптимизируют свое энергетическое состояние в предоставленном вместилище, точно так же биологические виды оптимизируют себя применительно к среде обитания. Морской леопард конструктивно явно менее совершенен, чем его сухопутный тезка, но к своей среде обитания приспособлен великолепно. И вообще, возврат водных млекопитающих в море с редукцией конечностей и целого ряда других "достижений" эволюции скорее заполнение тихой заводи, чем "стрежень" эволюции.
Неизменность среды обитания обеспечивает неизменность обитателей. Тому примеры латимерия, гаттерия и множество других "живых ископаемых". Изменения начинаются тогда, когда меняются свойства "вместилища" (например, меняется климат), либо "вместилище" переполняется и нужно осваивать новые места обитания. Поэтому на модный, но нелепый вопрос: "почему сейчас обезьяны не развиваются в людей?" следует ответить - "незачем, да и некуда". Скорее можно ожидать обратного превращения, некоторые свидетельства которому уже налицо. Нам представляется, что для биологии объективно настало время перейти от стадии накопления фактов к обработке и анализу накопленного.Материальное обеспечение перехода к стадии анализа.
Мы полагаем, что радикальных изменений не будет, но наверняка нас ожидает не одна сенсация. Например, для ярых противников родства с обезьянами может оказаться "утешением" то, что свинья ближе к нам, чем обезьяна, не только по аминокислотному составу белков и поведению "под дубом", но еще и "генеалогически".Собственно говоря, приступить к такому анализу можно было несколько десятилетий тому назад, но… Проблема заключалась в том, что не было понятно, каким образом линейная последовательность аминокислот, закодированная в НК и реализуемая при синтезе белков, обеспечивала и трехмерное развитие организма, и дифференциацию его клеток, и все другие многочисленные наследственные свойства. К счастью для биологов на рубеже веков появились фракталы и, тем самым, был создан математический аппарат, позволяющий не только понять механизмы реализации наследственной информации, но и научиться в обозримом будущем рассчитывать их.
Для примера приведем построение одного из простейших фракталов - фрактальной кривой, называемой драконом Хартера-Хейтуэя.
Пусть образующим элементом будут два равных отрезка, соединенных под прямым углом. В нулевом поколении единичный отрезок заменим этим образующим элементом так, чтобы угол был сверху. Можно сказать, что при такой замене происходит смещение середины звена. При построении следующих поколений выполняется правило: самое первое слева звено заменяется образующим элементом так, чтобы середина звена смещалась влево от направления движения, а при замене следующих звеньев, направления смещения середин отрезков должны чередоваться.Примерно по такому алгоритму строят свои дома термиты, - каждый рабочий термит строит глиняный столбик, влезает на него и соединяет вершину с другим ближайшим столбиком перемычкой, затем на перемычке строит новый столбик и т.д.
Легко понять, что в качестве образующего элемента может быть молекула белка с двумя не симметрично расположенными "точками связи". Если "точек связи" более двух, получаемые фигуры будут трехмерными, а их конфигурация будет разительно меняться при малейшем изменении взаимного расположения "точек связи". В свою очередь, расположение "точек связи" однозначно задано первичной структурой белковой молекулы, следовательно, может быть "записано" в НК.
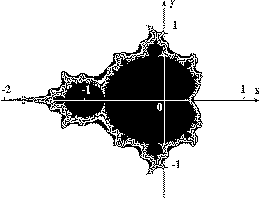

На рис. 2 показаны изображения простейших "алгебраических" фракталов, известных как множества Мандельброта. Алгоритм построения достаточно прост и основан на итеративном выражении: z[i+1] = z[i] *z[i] + c, где z[i] и c - комплексные.
Итак, в настоящее время в биологии есть огромнейший фактический материал, есть "чертежи", по которым работает живое производство, понятны элементарные "производственные процессы" и, наконец, появился математический аппарат для расчета "готовой продукции". Именно поэтому нам представляется, что "полевые" изыскания пора менять на "камеральную" обработку, сопровождаемую небольшим количеством радикальных экспериментов с точно поставленными задачами.Обновленная трактовка дарвиновской теории.
Тем более что первое успешное знакомство биологов (в лице ботаников) с фрактальными методами уже состоялось.
Знание механизма наследственности, наличие в хромосомах механизма репарации повреждений нитей НК вынуждает признать, что повреждения и сбои - явления достаточно частые и не только вполне обыкновенные, но и необходимые. Подавляющая их часть немедленно исправляется репарационными механизмами клетки и последствий не имеет. Не исправленные повреждения (например, разрывы в одном и том же месте обеих нитей двойной спирали или во время митоза в разделенных нитях) в большинстве своем приводят к прекращению синтеза и, в конце концов, к гибели клетки. И только очень малая часть таких повреждений оказывается совместимой с синтезом белка измененной структуры.Знание механизма синтеза белка позволяет понять, что большая часть изменений в первичной структуре белка не оказывает влияния на его работоспособность, лишь некоторые изменения структуры делают белок либо неработоспособным, либо работающим аномально.
В первом случае клетка рано или поздно гибнет, ликвидируя тем самым мутацию, и только аномально работающие белки выводят мутацию на надклеточный уровень. Но и это не все. Мутации в соматических клетках могут определить судьбу индивида, но не могут повлиять на генотип и, следовательно, на вид в целом. И только изменения в геноме клеток репродуктивного аппарата (и, возможно, стволовых клеток) могут быть переданы потомству и, тем самым, достичь уровня естественного дарвиновского отбора. Кстати, о стволовых клетках. Хотя их целевая функция пока до конца не ясна, можно предположить, что в многоклеточном организме они являются своего рода универсальными заготовками для регенерации, заменяя поврежденные специализированные и приобретая соответствующую специализацию "по месту". Действительно, для организма было бы расточительным как создавать резервы по каждой специализации, так и организовывать деление клеток в стадии глубокой дифференциации. Гораздо проще иметь (а при необходимости, и размножить) неспециализированный резерв, посредством которого и проводить соответствующие замены. Однако, если это так, то подобный механизм может служить своего рода транслятором мутаций и передавать их в репродуктивные клетки.Обратим внимание: мутации не вызываются изменением условий, - они происходят всегда.
При неизменных условиях организмы достаточно быстро (в течение нескольких поколений) выходят на оптимальное соответствие среде обитания, и новые мутации "гасятся" естественным путем на уровне вида. Путь несколько расточительный, но обеспечивающий достаточно быструю реакцию на изменение среды обитания. По сути, природа, задолго до математиков, "придумала" вариационный метод. Так, например, если бы при бурном распространении компьютеров в течение нескольких сотен лет в качестве органов управления сохранялись бы клавиатуры, для работы с которыми шестой палец явно полезен, число шестипалых операторов существенно бы увеличилось (именно шестипалых, а не пяти с четвертью, пяти с половиной и т.д.).Поэтому всякие рассуждения о "предваряющей" эволюции беспочвенны. Любой эволюционный шаг обязательно должен оптимизировать существование вида в измененной среде обитания. Именно вида как целого, мутации индивидуумов могут быть любые, включая летальные. Но ни один вид не может позволить себе "временное эволюционное отступление" в расчете на будущие блага.
Во избежание недоразумений при дальнейших построениях введем вспомогательную классификацию мутаций. Эта классификация не учитывает множества важных аспектов процесса (например, оставляет в стороне природу мутагенных факторов) и не претендует на самостоятельное существование. Итак, с чисто "технической" целью мы предлагаем различать следующие виды мутаций:Теперь можно скорректировать теорию эволюции Дарвина.
1. Мутации замещения. Как и следует из названия, такие мутации возникают при замещении в последовательности НК одних нуклеотидов другими, в результате чего изменяется кодируемая аминокислота. Замещения в пределах избыточности кода, не приводящие к смене аминокислоты, игнорируются.
2. Мутации рекомбинации. Репарационный аппарат клетки не в состоянии исправить одновременный разрыв сразу двух нитей НК. Существует отличная от нуля вероятность, что полученные при нескольких таких разрывах фрагменты смогут соединиться между собой в ином сочетании. Такие мутации мы обозначим как рекомбинационные.
3. Мутации прерывания. При нормальном функционировании клетки ее генетический аппарат активно участвует в двух процессах - в копировании генетической информации, при котором двойная спираль частично "расплетается" и на этих участках синтезируются матричные РНК, и митозе (делении клетки), при котором двойные спирали "расплетаются" полностью и на каждой из нитей синтезируется комплиментарная нить. Оба процесса, а особенно второй из них, весьма сложны и требуют четкой синхронизации всех операций. Внешнее воздействие (радиация или какой-то химический ингибитор) в это время могут либо прервать процесс, либо привести к пропуску какой-либо стадии. Результатом таких мутаций может быть увеличение числа хромосом вплоть до удвоения, если процесс прерван на одной из последних стадий митоза. Такие мутации обозначим как мутации прерывания.
Изменения вида начинаются при изменении условий обитания, к которым относятся и изменения климата, и дефицит кормовой базы, вызванный, например, чрезмерным размножением представителей самого вида, и появление либо размножение его фагов и тому подобное. С точки зрения эволюции и внутривидовая, и межвидовая борьба - всего лишь форма изменения условий обитания, и совершенно не существенно, не смог ли погибший индивид увернуться от хищника или от камня в зоне камнепада.В изменившихся условиях какая-то из мутаций обеспечивает лучшую приспособленность, в течение нескольких поколений становится доминантной и вид снова стабилизируется. Как правило, за приспособляемость вида ответственны, главным образом, мутации замещения. Такие мутации не препятствуют скрещиванию, напротив, перекрёстное оплодотворение способствует их закреплению.
Можно довольно основательно предполагать, что появление разветвленных хромосом является результатом мутаций рекомбинации, дающих обычно сразу несколько радикальных изменений фенотипа. Если такой индивидуум окажется более приспособленным к существующим условиям, мутация закрепится в потомстве и приведет к появлению подвида или новой породы. Здесь перекрёстное оплодотворение, как правило, возможно, но при этом зачастую происходит возврат потомства к прототипу.А вот за появление новых, существенно отличающихся от предшественников видов, ответственны мутации прерывания, довольно часто приводящие к увеличению числа хромосом, вплоть до удвоения.После нескольких рекомбинаций и замещений дублировавшие друг друга хромосомы дифференцируются и дают клетки с почти удвоенным потенциалом дальнейшего развития. Перекрёстное оплодотворение с прототипом невозможно, отмечены только весьма редкие случаи, когда сперма самца прототипа провоцирует "партеногенетическое" (на самом деле, мало понятное) развитие зародыша. Не исключено, что пресловутый "кембрийский взрыв" есть следствие одной или нескольких мутаций прерывания.
Каждый эволюционный шаг происходит достаточно быстро (скорее всего, экспоненциально) из-за возникновения положительной обратной связи – появление новых, более приспособленных особей ухудшает условия существования не изменившихся и стимулирует размножение измененных.Вторым важным дополнением теории эволюции является "правило оптимума" (назовем его так), то есть любой шаг эволюции вида, хотя бы незначительно, но приближает вид к оптимуму в текущих условиях среды обитания. Никакие изменения, хотя бы и сулящие огромнейший прогресс в будущем, но отдаляющие вид от оптимума в настоящем, невозможны. Более того, некоторые достижения, например, вскармливание потомства неким подобием молока, реализованное амазонскими рыбами дискусами и арапаимами (крупнейшая в наше время пресноводная рыба), могут отсутствовать у ближайших генетических родственников и проявиться у совсем далеких видов.
Как видим, предлагаемые поправки к теории эволюции Дарвина довольно радикальны, но отнюдь не уничтожают главное - естественный отбор - важнейшую обратную связь между видом и средой обитания.Поэтому нет оснований, подобно Уэссону, отбрасывать дарвиновскую эволюцию, как "потачку стародавней грезе о Вселенной, уподобленной огромному часовому механизму", заменяя её "эволюцией по Уэссону":
Неясно только, чем "Великий Хаос", всемогущий, вездесущий и целенаправленный, лучше "Великого Логоса", "огромного часового механизма", а также целого сонма всемогущих и вездесущих Богов и божков древних. Но это уже вопрос веры, а не науки.Нам кажется, что, во-первых, нет смысла "обожествлять" математический аппарат до "вселенской причины", а во-вторых, разрушать до основания если и не дворец, то еще довольно прочный дом, чтобы на его месте построить хлипкую хижину.
Читатель может сам убедиться в том, что теория эволюции Дарвина, дополненная принципом дискретности и принципом оптимума, становится внутренне непротиворечивой и всеобъемлющей. В особенности, если не забывать о том, что Земля непрерывно изменяется, что всего 200 млн. лет тому назад существовала Пангея, что Альпы совсем недавно были дном моря Тетис и что подобные перемены не могли не влиять на живые организмы.Эволюция Homo sapiens.
Действительно, переход к прямохождению с открыванием врагу самой незащищенной части – живота, оправдан только при наличии уже достаточно развитого мозга. В то же время развитие мозга без его немедленного применения невозможно, то есть здесь налицо то самое “тактическое отступление” в расчете на “светлое будущее”. Такие шаги в эволюции принципиально запретны. Отсюда неудачи палеонтологов в поисках “недостающего звена”. Трудно найти черную кошку в темной комнате, особенно, если ее там нет, и никогда не было.Для того чтобы поиски стали успешными, необходимо вести их на таком пути эволюции приматов, на котором каждый эволюционный шаг давал бы преимущество в борьбе за выживание. Как нам кажется, именно такой путь, встречая яростное сопротивление и дарвинистов, и их антиподов, в течение двух десятилетий прорабатывает российский ученый Л. И. Ибраев [1].
Он предлагает искать предков человека среди приматов, обитавших в прибрежных лесах типа теперешних мангров. В настоящее время в мангровых лесах Калимантана обитают приматы “носачи”, которые значительную часть времени проводят в воде. Эти обезьяны великолепно владеют прямохождением, хотя их мозг гораздо примитивнее мозга шимпанзе, волосяной покров у них развит только на голове, зато, как и у человека, развит слой подкожной жировой клетчатки, пот содержит много соли и т.п. Руки носачей хорошо развиты и универсальны, даже процесс зачатия у них аналогичен человеческому – лицом к лицу, что характерно для водных млекопитающих, но не встречается у сухопутных, в том числе, и человекообразных.Впрочем, дадим слово автору идеи:
"Мне думается, многие противоречия и неясности (антропогенеза, ВК) устраняются выдвинутой мною в 1985 г. теорией: непосредственными предками людей были не волосатые, маломозглые и неуклюжие выходцы из леса, как предполагал Ч. Дарвин, а голые большеголовые и двуногие прибрежные обезьяны. Назовем их (по имени древнегреческих речных нимф - наяд) наяпитеками (Pithecus naias). Они обитали в неогене по берегам рек, ручьев, озер и других пресноводных водоемов в полусаванной предгорной местности. Питались ловлей и собиранием моллюсков, раков, лягушек, черепах, грызунов, птичьих яиц, прибрежных ягод, фруктов и других плодов, корений и насекомых и использовали для ловли и вскрытия раковин и панцирей расколотую гальку, палки и кости. Прежнее древесное существование, развившее у них гибкие и цепкие пятипалые лапы, цветное бинокулярное зрение, исключительную пространственную координацию движений, увеличенную затылочную зрительную и теменную кинестетическую кору головного мозга, стало быть, сообразительность, - хорошо подготовило их к такому способу прибрежной жизни, которую не ведет ни одно другое животное. <…>Гипотеза Ибраева полностью соответствует "принципу оптимума" - каждый эволюционный шаг дает виду немедленный выигрыш, поэтому предлагаемые им в качестве человеческих пращуров "наяпитеки", сходные с современными носачами, заслуживают самого пристального внимания.
Вода и занятость передних лап воспрепятствовали наяпитекам опуститься на четвереньки и обусловили развитие прямохождения. Дно мелководий, часто мягкое, потребовало больших плоских ступней. Полуводное существование под палящим тропическим солнцем, от которого уже не защищал лес, и переохлаждение от мокрой шерсти в тени привели к утрате наяпитеками шерстяного покрова. На темени же, реже окунаемом в воду и особенно страдающем от солнца, волосы, наоборот, выросли. <…>
Оголение кожи у наяпитеков сопровождалось развитием слоя подкожного жира, хотя и не столь толстого, как у свиней, бегемотов, носорогов и других полуводных млекопитающих, обитающих в жарком климате. Кроме того, увеличилось количество потовых желез (до двух-пяти миллионов), что также спасало наяпитеков от перегрева. <…>
Отношение длины кишечника к длине тела у человека равно 5,6, что соответствует питанию его предков преимущественно моллюсками и ракообразными, находясь как раз между такими показателями рыбоядных (4,5) и всеядных (6,8), но далеко отстоит от плотоядных (3,7), зерноядных (8,7) и тем более травоядных (15,1)."
Легко видеть, что переход к прямохождению позволял им расширить зону добычи пищи, в то же время нападать на незащищенный живот в их среде обитания было просто некому.Майкл Бейджент в своей во многом (и порой очень многом) спорной книге "Запретная археология" указывает еще на одно сходство между носачами и людьми - “опущенную гортань”, хорошо известную биологам особенность человека, отличающую его от всех сухопутных животных (и чуть ли не акт “божественного творения” по мнению клерикалов).
У всех сухопутных млекопитающих, кроме человека, дыхательный тракт отделен от пищевого канала. Верхняя часть трахеи - гортань - расположена над мягким нёбом, в котором есть соединительное отверстие, постоянно закрытое кольцевой запирательной мышцей - сфинктером. В необходимых случаях, например, для издания звуков, сфинктер расслабляется, гортань опускается в ротовую полость, и каналы сообщаются между собой. По истечении надобности сфинктер снова запирается и разобщает каналы.
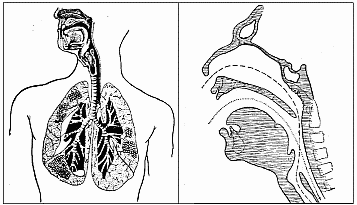
Это существенно осложняет нам жизнь, превращая глотание в сложную, управляемую мозгом процедуру. Но именно так устроено горло не только у людей, но и у тюленей, дельфинов, дюгоней и китов, то есть у водных млекопитающих (и у носачей). Короче говоря, у тех, кому требуется сделать мощный быстрый вдох перед погружением или после погружения в воду.Видимо, именно в этом эволюционный смысл "опущенной гортани". Кроме того, что этот факт подтверждает гипотезу Ибраева о "болотном", а не "саванном" происхождении человека, именно такое устройство горла с постоянным его контролем со стороны высших отделов мозга обеспечило богатство акустических возможностей человека и, главное, членораздельную речь.
Таким образом, если принять гипотезу Ибраева, предки человека - наяпитеки - обладали физиологической возможностью богатой акустической артикуляции, способной обеспечить членораздельную речь задолго до соответствующего развития головного мозга, и получили её “бесплатно”, в качестве приложения к “болотному” образу жизни. Это позволяет сделать предположение о совершенно ином пути развития интеллекта у наших далеких предков.Эволюция разума.
Признак хорош, прежде всего, тем, что, как правило, орудия труда наши предки изготавливали из камня или костей, материалов весьма долговечных и стойких, способных сохраняться десятки и сотни тысяч лет и "доживать" до рук археологов. Но этот подход несколько напоминает поиск под фонарем потерянного в темноте кошелька только потому, что под фонарем светло.Действительно, орудия труда используют очень многие животные различных видов, - от птиц до млекопитающих. Причем не только используют разово, но и хранят, и даже изготавливают! (Например, обламывают колючку для добывания червей до оптимальной длины). Тем не менее, никому не придет в голову считать дельфинов, не использующих орудий, глупее вороны. Видимо, выбран не тот критерий.
Зоопсихологи (показательно, кстати, и то, что они появились) хорошо знают, что животные не только инстинктами сильны, но и думать умеют, и действия свои планируют. И что родители обучают своих детей, в том числе, и личному опыту. Однако в отличие от инстинктов, личный опыт животные могут передавать только показом, на уровне первой сигнальной системы. А это налагает большие ограничения и на скорость, и на объем переданной информации, а самое главное, очень велики потери.А вот побочный эффект "опущенной гортани", резкое увеличение возможностей акустического общения (рис. 4) привело, по нашему мнению, к появлению второй сигнальной системы и резкому увеличению объема передаваемого потомству личного опыта благодаря “акустическому сопровождению”.
Естественно, резкий "интеллектуальный" рывок не мог не отразиться на первобытном "производстве", следовательно, по времени появление второй сигнальной системы и массовое изготовление орудий труда должны практически совпадать.Так что действительно "вначале было слово". Окончательное же закрепление человечества в "интеллектуальной среде обитания" произошло с появлением письменности - пиктограмм, рун, иероглифов и т.п., что и привело к абстрактным понятиям, математике и вообще науке.
Однако если принять такой путь эволюции человека разумного, то приходится признать и некоторые неприятные следствия. Выход человека в информационное измерение в значительной мере вывел человека из-под власти естественного физиологического отбора. Он получил возможность, не меняясь генетически, повышать свои потребительские возможности. Для того чтобы обычный хищник заметно повысил свои возможности ловли добычи, ему нужно измениться самому. Параллельно будет совершенствоваться и добыча, поэтому равновесие не будет нарушено.Иное дело человек. Сам не изменяясь, он совершенствует свои орудия, что намного быстрее естественного развития любого вида, и нарушает природное равновесие в свою пользу. Как и следует ожидать, такое нарушение не проходит даром, люди начинают брать от природы много больше необходимого, "жиреют" в прямом и переносном смысле и … вырождаются. Вполне допустимо, что Римская империя, Древний Египет, Атлантида (если она была) были лишь последними из рухнувших цивилизаций. Возможно, цивилизации возникали и рушились и двадцать, и сорок тысячелетий тому назад, возможно, легендарный "золотой век" - всего лишь начало применения металлов в человеческом быту, ибо золото не нужно выплавлять.
На наш взгляд, сейчас человеческая цивилизация успешно приближается к очередному коллапсу, если не сделает должных выводов из собственной истории и не станет заботиться о питании разума больше, чем о заполнении желудка в широком смысле слова.Суть именно в том, что материальное потребление человека изменяется практически независимо от его физиологических изменений. Точнее, в том, что биосфера не успевает реагировать на человека эволюционно, поэтому реагирует революционно - разрушением.
Обычно появление и последующая динамика нового вида происходит так, как показано на рисунке 5 сплошной линией.
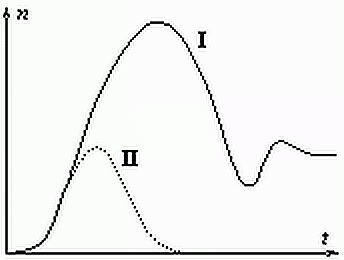
Рис. 5. Динамика развития
нового вида во времени после его появления (I). Пунктиром (II) показана
возможная динамика вида Homo sapiens.
На первых порах влияние нового вида на окружающую среду ничтожно, конкурентов нет, "фаги" его еще "не распробовали", вид размножается быстро и без помех. Но по мере роста численности вида растет и его воздействие на окружающий биоценоз: заметно начинает уменьшаться численность биообъектов, служащих субстратом новому виду (например, редеет любимая трава или уменьшается количество зайцев и т.п.), зато растет количество биопродуктов - результатов существования нового вида.Иначе говоря, вид начинает влиять на среду своего обитания. Эти изменения запускают эволюцию всего биоценоза в ареале нового вида: появляются специфические для нового вида вирусные и микробные болезни, хищники (которым теперь стало труднее ловить зайцев) заносят его в разряд добычи, да и сам вид испытывает все большие трудности и в пропитании, и в размножении. Наконец, антагонистические процессы останавливают прирост, начинается сокращение, так как процесс обладает положительной обратной связью.
Обратим внимание: биосфера реагирует не на численность нового вида, а на вызванные им изменения среды обитания, от величины этих изменений зависит диапазон и величина реакции биоценоза. Для всего живого, кроме человека, численность вида и его влияние на среду обитания практически синонимы, поэтому реакция биоценоза оказывается адекватной и после нескольких затухающих колебаний численность нового вида стабилизируется на оптимальном уровне.С человеком не так. Мало того, что благодаря разуму он все более успешно противостоит естественным антагонистам (в том числе, бактериям и вирусам), он исчерпывает продуктивность своей экологической ниши задолго до достижения критической численности. Так, в настоящее время один житель "индустриально развитых" стран (читай, общества потребления) потребляет столько же ресурсов, сколько 20 человек из "развивающихся". Неудивительно, что реакция биосферы оказывается существенно большей, чем это допускает реальная численность. А вот в этом случае процесс может (хотя отнюдь не обязан) пойти и так, как показано на рис. 5 пунктиром. То есть не к оптимуму, а к уничтожению.
Возможно, именно это и происходило не раз в истории человечества с исчезнувшими цивилизациями. Но тогда безудержный рост потребления носил локальный (региональный) характер, природные защитные механизмы тем или иным путем ликвидировали очаг опасности, оставляя нетронутыми менее "цивилизованные" сообщества. Иное дело сейчас. Психология потребления приобрела практически глобальный статус, поэтому, скорее всего, и ответные шаги биосферы будут адекватны.
<…>"за свое последнее столетие обитатели Надежды потеряли контроль над развитием технологии и практически необратимо нарушили экологическое равновесие. Природа была уничтожена. Отходы промышленности, отходы безумных и отчаянных экспериментов в попытках исправить положение загадили планету до такой степени, что местное человечество, пораженное целым комплексом генетических заболеваний, обречено было на полное одичание и неизбежное вымирание.
<…>Внешне это выглядело как стремительное, нелинейное во времени ускорение темпов развития всякого мало-мальски сложного организма. Если говорить о человеке, то до двенадцати лет он развивался, в общем, нормально, а затем начинал стремительно взрослеть и ещё более стремительно стареть. В шестнадцать лет он выглядел тридцатипятилетним, а в девятнадцать, как правило, умирал от старости".
Прогерия взрослых, или синдром Вернера описан впервые еще в 1904 г. Страдающие им люди развиваются с нормальной скоростью до 17–18 лет, а потом начинают стремительно стареть. Лишь немногие дотягивают до пятидесяти, уходя из жизни глубокими стариками. У них быстро развивается широкий спектр всевозможных патологий, обычно связываемых с возрастными изменениями – атеросклероз, диабет, катаракта, различные типы доброкачественных и злокачественных опухолей. В Японии частота этого заболевания существенно выше, чем в других странах, и достигает одного случая на сорок тысяч.В результате генетического анализа удалось выяснить, что прогерия взрослых является аутосомным рецессивным заболеванием. Это означает, что оно проявится в зрелом возрасте только у тех новорожденных, которые получили одновременно от каждого из родителей по определенному мутантному гену, находящемуся в восьмой хромосоме.
Наиболее трагично протекает прогерия детей, которую называют еще синдромом Хатчинсона–Гилфорда. Ребятишки с этим страшным диагнозом стремительно стареют. В среднем они едва дотягивают до 12 лет и чаще всего умирают в этом юном возрасте от банальных старческих инфарктов. К этому времени они и выглядят как глубокие старики – лысеют, страдают от атеросклероза и фиброза миокарда, практически полностью лишаются подкожного жирового слоя, теряют зубы... К счастью, рождаются такие дети чрезвычайно редко, с частотой один на миллион.
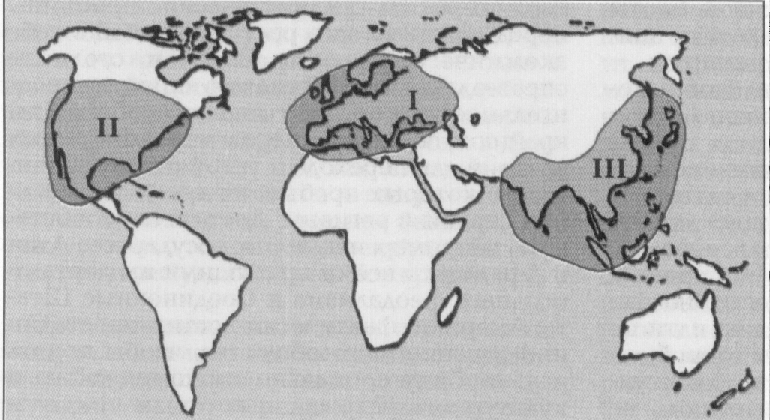
С точки зрения сохранности природных экосистем все страны мира могут быть сгруппированы по трем категориям:страны с хорошими стартовыми условиями перехода к устойчивому развитию (площадь ненарушенных экосистем превышает 60% их территории) - 8 стран, или 5,5% от всех государств мира; страны с промежуточными стартовыми условиями (от 59% до 10% сохранившихся природных территорий) - 47 стран, или 32,5% общего числа; страны с низкими стартово-экологическими условиями, практически не способные к реставрации (менее 10% площади сохранных экосистем) - 91 страна, или 62% от всех государственных образований.
Подобные примеры, на наш взгляд, делают "сценарий Стругацких" более чем реальным. Более того, генные инженеры уже испытывают на мышах вирусы, которые должны "исправлять" геном человека. Имеется в виду, прежде всего, лечение прогерий и добавление теломеров с целью продления жизни.
Между тем, существует вполне разумный (именно, разумный) путь - направить потребление человека в область, где у него нет не только конкурентов, но и каких-либо ограничений, - в область питания разума информацией. Разумеется, ограничив строго необходимым физиологическим минимумом материальные потребности - нечто подобное йоге, но на активном, а не умозрительном уровне. Примерно так, как предлагал П.П. Ветров в своей статье "Разумен ли Homo sapiens?" ("Виртуальный мир" N 1 2003 г.).
Понятно, что человечество, не сумевшее не злоупотребить второй сигнальной системой, тем более не пригодно к освоению третьей.
1. Эволюция - принципиально дискретный процесс, как в структурном, так и во временном отношении, основанный на частом появлении и "гашении" мутаций.Литература
2. Время приспособления вида к изменившимся условиям составляет всего несколько поколений, поэтому плавное изменение условий отслеживается практически без запаздывания.
3. Каждый шаг эволюции вида приближает вид к оптимуму, обратный процесс невозможен.
4. Подлинным "питеканторопом", видимо, был "наяпитек".
5. Первичным признаком зарождения интеллекта является вторая сигнальная система, изготовление же орудий труда - вторично.
6. Ближайшей жизненно важной задачей человека является "переключение" своих потребностей из материальной в духовную сферу.