М.В.Листов (д.б.н.)
А.И.Мамыкин (д.ф-м.н.)
Запись на плазмалемме*.
Фейк или реальность?
Диффузионный поток отрицательно заряженных свободных радикалов и перенос
неспаренного электрона супероксидом и другими свободными радикалами являются
составляющими энергетического обмена в биосистеме и условием активного
состояния электромагнитного поля в структурах головного мозга, трофической
функции нервных волокон, покрытых миелином. Важная роль активных форм кислорода
(АФК) в физиологических процессах, происходящих в организме, во многих
случаях обусловлена действием потока активных частиц отрицательно заряженных
радикалов супероксида из жидких субстанций организма на микроструктуры
наружной поверхности клеточной мембраны. В структуре интенсивно исследуемого
биофизиками бислоя липидов мембраны гидрофильные полярные головки молекул
липидов (и холестерина) внешнего и внутреннего слоёв мембраны ориентированы
к водной фазе вне и внутри клетки под разными углами по причине подвижности
отдельных молекул фосфолипидов в плоскости клеточной поверхности и их вращения
в плоскости, перпендикулярной плазмалемме.
Липопротеиновые фрагменты липидов, холестерина и структурных белков с ионофильными
полюсами соединены между собой межмолекулярными силами взаимодействия в
кластеры и плоты, площадь которых в сумме доходит до 20 % от общей клеточной
поверхности, причём заряд этих участков отрицателен. Отдельные кластеры
заякорены с помощью белковых нитей так называемого субмембранного комплекса
поверхностного аппарата клетки, другие, их называют плотами, имеют свободу
латерального перемещения по поверхности. Одна из возможных схем распределения
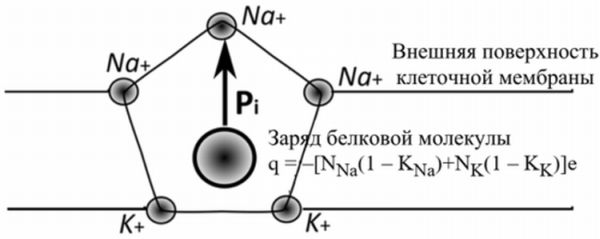
зарядов в активном центре плазмалеммы клетки представлена на рисунке. Плоскостная
модель (легко достроить до объёмной структуры) активного центра в форме
пентагона состоит из отрицательно заряженного ядра, молекулы структурного
белка, заряд которого компенсирован зарядом ионов Na+, K+ и аквакомплексов
(Na.6H2O
и K.4H2O)
на внешней и внутренней поверхностях мембраны.
 Плазмалемма исходно имеет гидрофобную поверхность, которая в результате
внедрения активных белковых центров и дефектов структуры становится гидрофильной
в результате уменьшения свободной поверхностной энергии. Существенным для
понимания механизма взаимодействия поверхности клеточной мембраны с диффузионным
приповерхностным слоем является то, что поверхностные группы выступают
в роли центров адсорбции, поскольку атомы водорода, принадлежащие сольватным
оболочкам ионов Na+ и K+, способны образовывать водородные связи, координируя
неподелённые электронные пары кислорода сорбированных из диффузионного
слоя молекул воды с образованием связанных ионов гидроксония Н3О+,
которые и формируют двойной электрический слой дипольной сети плазмалеммы.
Близкие по энергетике процессы происходят на поверхности пористого кремния,
где роль активных центров играют Si-Hx группы и наноразмерные поры.
Плазмалемма исходно имеет гидрофобную поверхность, которая в результате
внедрения активных белковых центров и дефектов структуры становится гидрофильной
в результате уменьшения свободной поверхностной энергии. Существенным для
понимания механизма взаимодействия поверхности клеточной мембраны с диффузионным
приповерхностным слоем является то, что поверхностные группы выступают
в роли центров адсорбции, поскольку атомы водорода, принадлежащие сольватным
оболочкам ионов Na+ и K+, способны образовывать водородные связи, координируя
неподелённые электронные пары кислорода сорбированных из диффузионного
слоя молекул воды с образованием связанных ионов гидроксония Н3О+,
которые и формируют двойной электрический слой дипольной сети плазмалеммы.
Близкие по энергетике процессы происходят на поверхности пористого кремния,
где роль активных центров играют Si-Hx группы и наноразмерные поры.
Энергетическая неоднородность поверхности клеточной мембраны определяется
прежде всего различным строением металла-координатора, а также ассоциированных
молекул воды и аквакомплексов противоионов, в зависимости от того, формируются
они в виде отдельных кластеров вокруг отрицательно заряженного ядра молекулы
структурного белка (см. рисунок) и адсорбированных им ионов металла, или
покрывают фрагмент плазмалеммы с дефектами билипидного слоя. В первом случае
аквакомплексы формируются как с внешней, так и с внутренней поверхности
плазмалеммы, тогда как для компенсации отрицательного заряда дефектной
структуры липидного бислоя достаточно кластера ассоциированных молекул
Н2О с его внешней стороны.
Ещё одно важное различие состоит в том, что аквакомплексы, образующиеся
вокруг интегрированных в билипидный слой структурных белков, формируют
кластеры под действием центральной силы притяжения к молекуле белка, которая
убывает с увеличением расстояния медленнее сил отталкивания. Радиус действия
межмолекулярных сил, напомним, не превышает 10 нм и распространяется на
структуры с пониженной размерностью, в том числе на вторичные и третичные
структуры гетерополимеров многоклеточной биосистемы, белков и нуклеиновых
кислот, а также ДНК и РНК вирусов, неклеточных паразитов растений, животных
и человека.
Адсорбция противоионов дефектных участков плазмалеммы клеток ядерных организмов
в свою очередь происходит под действием сил, равномерно распределённых
по участку с нарушением структуры бислоя молекул липидов и липоидов, быстро
спадая по мере удаления от границ дефектного участка. Подобная неоднородность
поверхности приводит, как правило, к формированию фрактальных структур,
которые обладают свойством памяти.
Определенный вклад в формирование
потенциала действия вносят свободные, т. е. некоординированные молекулы
воды, которые имеют большую свободу вращательных, возможно, поступательных
движений, а при ассоциации диполей образуются олигомеры молекул Н2О
за счёт водородных связей (Н2О)2,
(Н2О)3
с общей формулой (Н2О)n.
Предполагают, что при 0 °С вода состоит в значительной мере из молекул
(Н2О)3,
при нагревании до 4 °С утроенные молекулы диссоциируют с образованием удвоенных
молекул (Н2О)2,
которые сообщают воде повышенную плотность, вероятно из-за наличия в спаренных
молекулах двух водородных связей. Принимая во внимание, что молекулы воды
водородными связями могут объединяются в цепочки (Н2О)n
и сети, можно предположить значительную роль молекул воды и её комплексов
с металлами в поддержке кооперативных процессов, таких как поляризация
мембран с передачей заряда через эстафетный механизм движения протонов
по системе водородных связей, или связывание молекулы кислорода специальными
белками-переносчиками крови, зависящее от рН. Водные системы (кровь, лимфа,
ликвор, межклеточная жидкость) находятся в основе жидкой внутренней среды
организма, содержащей супероксид, фактор, влияющий на плазмалемму и ее
потенциал.
Вода является основой биосистемы, воду содержат также липопротеиновые клеточные
мембраны организма, а дипольная сеть биосистемы включает протяженные и
координированные структуры, обладающие пониженной размерностью. Клетка
является структурной, функциональной и генетической единицей организма,
её внешняя мембрана служит основой дипольной сети биосистемы.Существенно,
что на липопротеиновой матрице клеток происходит как локальная деполяризация,
так и образование пористой структуры в результате окисления поверхностных
центров отрицательно заряженными радикалами, перенос которых на поверхность
осуществляется диффузионным потоком анион-радикалов кислорода, при этом
изменение плотности потока АФК неизбежно модулирует фрактальную структуру
кластеров на плазмалемме.
Энергетическая неоднородность поверхности клеточной мембраны обусловлена
прежде всего различным строением противоионных аквакомплексов, в зависимости
от того, формируются ли они в виде кластеров вокруг отрицательно заряженных
интегрированных белков или покрывают часть плазмалеммы с дефектами структуры
билипидного слоя. Таким образом, характер поляризации плазмалеммы, а следовательно
форма и интенсивность потенциала действия на ней, существенно зависят от
топологии и поверхностной концентрации интегрированных белков, а также
от формы и расположения дефектных участков плазмалеммы, определяя характерное
поведение импульсов действия для эукариотических клеток различных органов,
тканей и других составляющих биосистемы.
Межмолекулярные взаимодействия (в том числе непосредственно вокруг репликационной
вилки) происходят при сближении частиц и попадания в зону действия сил
Ван-дер-Ваальса с радиусом действия до 10 нм. Величина сил может изменяться
и зависит, например, от агрегатного состояния молекул воды. Явление сцепления
молекул воды и слипание клеток при адгезии бактерий на поверхности фагоцита
напоминают взаимодействия на уровне атомов и молекул. Молекулы ДНК располагаются
на некотором равновесном расстоянии друг от друга, обеспечивая тем самым
масштабную инвариантность и стабильность нуклеотидных последовательностей
генетической памяти.
Метилирование по цитозину с помощью ДНК-метилтрасфераз изменяет расстояние
между нуклеотидами, что блокирует экспрессию генов сайта, укрытого под
импринтом. Внутриклеточные объемы (компартменты), ограниченные липопротеиновой
мембраной везикулы и цистерны мембранных органелл, также являются объектами
с пониженной размерностью. Само ядро клетки, по сути, двухмембранный органоид,
внешняя мембрана которого связана с плазмалеммой и внутренней жидкой средой
каналами. Самособирающийся комплекс поверхностного аппарата живой клетки,
синтезируемые клеткой молекулы билипидного слоя и структурные биполярные
белки образуют дипольную сеть тканей и органов.
Клетки реагируют на местное и общее повышение концентрации свободных радикалов
во внутренней среде организма изменением топологии фрактальных поверхностных
структур, ростом частоты локальных деполяризаций и осцилляций плазмалеммы,
которая на большей части поверхности клетки представлена липидным доменом
с холестерином и структурными белками, пронизывающими билипидный слой.
Токи смещения, возникающие в процессе одноэлектронного переноса радикалами,
приобретают синфазный характер, подобно вынужденным переходам в квантовых
системах, в результате чего формируются импульсы солитонного типа, которые
в сетях нейронов передаются в биосистеме на большие расстояния.
Фрактальная поверхностная структура на плазмалемме, например, нервной клетки
формирует локальный дипольный момент интегрированной в липидный бислой
мембраны терминальной части молекулы структурного белка (его полярного
полюса) и дипольных моментов, создаваемых зарядами адсорбированных ионов
и ассоциированных в комплекс с ними молекул воды. При этом квантование
происходит только в одном направлении (поперечном), тогда как вдоль канала
квантования не происходит, поэтому электромагнитный импульс, записанный
во фрактальной структуре, может быть объединён в нейронную сеть.
* Плазмалемма (клеточная
мембрана) – поверхностная, периферическая структура, окружающая протоплазму
растительных и животных клеток.
 В
оглавление
В
оглавление