М.В.Листов, А.И.Мамыкин
Активация
биомембран
Одноэлектронный
механизм блокировки и проведения нервного импульса
Большая
часть социально значимых заболеваний человека (болезнь Альцгеймера, ишемия
миокарда, инсульт, рак и др.), а также ряд патологических состояний, вызванных
ионизирующей радиацией и стресс-факторами, имеют свободно-радикальную этиологию
с одноэлектронным переносом, который и является механизмом патогенетического
действия свободных радикалов [1].
Во всех случаях при моделировании указанных патологий на мышах линии DВА/2
внутрибрюшинной инъекцией водного раствора тетракалиевой соли копропорфирина
III (копропорфирина) с активацией растворами солей металлов-координаторов
[1-5], у животных в первые минуты после инъекций наблюдалось судорожное
состояние, сходное с опистотонусом человека. Мыши, образовав плотную группу,
стояли на задних лапах 5-10 минут в судорожной позе, выход из которой сопровождался
медленным поочередным вытягиванием лап, а также спины и шеи.
Описываемое поведение мышей коррелирует с кинетикой свободных радикалов
в организме [2], что подтверждается регистрацией в сыворотке крови животных
гидроксильного радикала [3], являющегося конечным продуктом переноса электрона
[4]. Полученная модель опистотонуса позволяет в известной степени исследовать
физическую природу явления.
Копропорфирин, структура которого представляет собой тетрапиррольное кольцо
с чередующимися двойными и одинарными связями, является аналогом гема,
повышенная концентрация которого, так или иначе, связана с опистотонусом,
который у человека известен, как один из основных симптомов столбняка.
Опистотонус, наряду с другими неврологическими симптомами, отмечается при
синдроме Криглера-Найяра I типа, или наследственной гипербилирубинемии.
Таким образом, наши опыты на мышах хорошо проецируются на рассмотренные
неврологические ситуации.
Состояния по типу опистотонуса, а также тремор или нистагм возникают при
перевозбуждении групп нейромоторных единиц, которое может быть вызвано
блокировкой прохождения нервного импульса при переносе электрона через
клеточную мембрану. Ведущую роль в переносе электрона, скорее всего, играет
транспорт супероксидного иона, сопровождающийся переворотом полярной молекулы
интегрального белка клеточной мембраны. Для оценки структуры двойного электрического
слоя мембраны с разностью потенциалов около 70 мВ, достаточно рассмотреть
систему параллельных электрических диполей, интегрированных в билипидный
слой, среднее расстояние между которыми составляет 50 – 60 нм. Таким образом,
в пределах нейромоторной единицы формируется сеть электрических диполей
с межмолекулярной корреляцией, где в создании зарядов на поверхности клеточной
мембраны участвуют как молекулы связанной воды [6], так и сорбированные
интегральными белками ионы Na+
и K+.
Перенос электрона происходит следующим образом. Отрицательно заряженный
ион-радикал супероксида притягивается положительным зарядом внешнего слоя
клеточной мембраны и сорбируется на молекуле белка, частично компенсируя
положительный заряд диполя. Отрицательный заряд диполя, находящийся на
внутренней поверхности мембраны, притягивается положительными зарядами
внешнего слоя, что приводит к структурной конформации молекулы интегрального
белка с одновременным переворотом молекулы и её дипольного момента. Этот
механизм по-новому объясняет как перенос электрона через мембрану, так
и встречный перенос ионов Na+
и K+. Оценка энергии
перехода даёт значение около 0,5 ккал/моль.
Далее, супероксидный ион отторгается отрицательным зарядом внутреннего
слоя мембраны и перенос электрона на этом завершается. В области, где произошел
переворот дипольного момента, на поверхности мембраны возникает область
с зарядом противоположного знака, который стимулирует вынужденный переворот
соседних диполей. При достижении критического уровня деполяризации
– 50 мВ
– происходит генерация
нервного импульса.
Математическая модель этого физиологического процесса строится в предположении
изоморфности коллективных состояний микрочастиц в кристаллической решётке
твёрдого тела и молекул интегральных белков мембраны. Это дает возможность
использовать для анализа процессов в клеточной мембране распространённые
теоретические подходы, например, модель Хаббарда [7] и модель Андерсона
[8]. При этом фазовый переход мембраны, сопровождающийся изменением её
симметрии, электрических свойств и характера прохождения нервного импульса
[9], обнаруживает качественное сходство с переходами типа «антиферромагнетик
– парамагнетик – ферромагнетик», которые описываются t-J моделью [10].
В мембране коллективные свойства проявляет двойной электрический слой полярных
молекул белков с двумя предельными упорядоченными состояниями. В одном
из них все диполи ориентированы параллельно, в противоположном состоянии
ориентация соседних диполей антипараллельна. При параллельной ориентации
энергия мембраны максимальна и поддерживается работой АТФ. Минимум энергии
соответствует антипараллельной ориентации соседних диполей, однако, в этом
состоянии мембрана не проводит нервный импульс. Гомеостаз устанавливается,
как динамическое равновесие между спонтанными и вынужденными переворотами
диполей и ориентирующим действием АТФ.
Несколько видоизмененный гамильтониан Хаббарда в терминах нашей модели
имеет вид
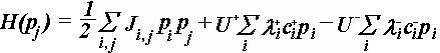 (1)
где индексы i, j нумеруют
узлы решётки (элементы белковой мозаики); pi
– дискретная «спиновая»
переменная, принимающая значения ±1; Jij
– энергия кулоновского
взаимодействия диполей, находящихся в узлах i, j; U+(-)
– энергия перехода
«спина» в параллельное (+1), или антипараллельное (-1) положение; ?+(-)
- соответствующие матричные элементы перехода; c+(-) - операторы повышения
(понижения). Верхний предел сумм определяется выбором числа ближайших соседей.
Гамильтониан (1) нелинеен, тем не менее, численный расчет позволяет построить
примерную фазовую диаграмму системы (рис. 1), на которой выделяются области
с различным типом взаимной ориентации диполей.
(1)
где индексы i, j нумеруют
узлы решётки (элементы белковой мозаики); pi
– дискретная «спиновая»
переменная, принимающая значения ±1; Jij
– энергия кулоновского
взаимодействия диполей, находящихся в узлах i, j; U+(-)
– энергия перехода
«спина» в параллельное (+1), или антипараллельное (-1) положение; ?+(-)
- соответствующие матричные элементы перехода; c+(-) - операторы повышения
(понижения). Верхний предел сумм определяется выбором числа ближайших соседей.
Гамильтониан (1) нелинеен, тем не менее, численный расчет позволяет построить
примерную фазовую диаграмму системы (рис. 1), на которой выделяются области
с различным типом взаимной ориентации диполей.
Кинетическое уравнение вытекает из (1) с учетом результатов [2]. Легко
видеть, что перевороты диполей также описываются нелинейным уравнением
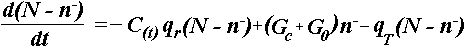 (2)
где N – полное число молекул
интегрального белка; n-
– число перевернутых
диполей; C(t) – зависящая от времени концентрация ион-радикала; G0
– скорость генерации АТФ при гомеостазе; GC
– зависящая от концентрации радикалов скорость генерации АТФ; qr
– вероятность переворота диполя под действием ион-радикала; qT
– термически равновесная вероятность спонтанного переворота полярной молекулы
белка.
(2)
где N – полное число молекул
интегрального белка; n-
– число перевернутых
диполей; C(t) – зависящая от времени концентрация ион-радикала; G0
– скорость генерации АТФ при гомеостазе; GC
– зависящая от концентрации радикалов скорость генерации АТФ; qr
– вероятность переворота диполя под действием ион-радикала; qT
– термически равновесная вероятность спонтанного переворота полярной молекулы
белка.
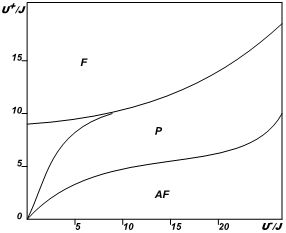
Рис. 1. Фазовая диаграмма
белковой мозаики мембраны.
F – область преимущественно
параллельной ориентации диполей; AF - область преимущественно антипараллельной
ориентации диполей; P–область распространения нервного импульса на фоне
помех
Уровень блокировки прохождения нервного импульса можно найти из порога
перколяции, значение которого здесь составляет около 0,45, что в пересчете
на концентрацию радикала в сыворотке крови дает 10-6
– 10-5 моль/л. Зависимость
концентрации радикала, рассчитанная на основе экспериментов [3] и кинетической
модели [2], представлена на рис. 2, где указан интервал существования опистотонуса,
который составляет около 7 минут.
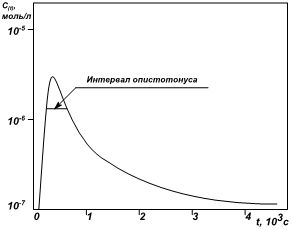
Рис. 2. Временная зависимость
концентрации супероксида в сыворотке крови (расчётная кривая)
Таким образом, нами получена экспериментальная модель опистотонуса, показывающая,
что этот неврологический симптом, имеет свободно-радикальную этиологию.
Теоретическое описание неврологических процессов проведено на масштабе
молекулярных взаимодействий и показано, что физиологические события в клеточных
мембранах изоморфны многочастичным квантовым явлениям в кристаллических
твёрдых телах. Последнее позволяет начать работу по созданию новых физических
методов регистрации энергетического спектра цепных реакций радикалов в
целях ранней диагностики многих заболеваний человека и их предотвращения.
Литература
-
Листов М.В., Мамыкин А.И. Фото- и химическая
активация одноэлектронного переноса при моделировании патологий на мышах
линии DBA/2. Доклады Академии наук. т.439, №6, 2011, с.841-843
-
Листов М.В., Мамыкин А.И. Кинетика релаксации
свободных радикалов и перенос электрона в жидких субстанциях организма.
Известия СПб ГЭТУ «ЛЭТИ», СПб, №3, 2010, с. 55 – 60
-
Листов М.В., Торопов Д.К., Родионов Г.Г.
Экспериментальное обоснование свободно-радикальной этиологии системных
заболеваний соединительной ткани на моделях полимиозита и гемозависимого
экзофтальма. Изд-во АН; ж. Докл. АН, т.414, №5, 2007, с.715-717.
-
Листов М.В., Мамыкин А.И. Эффективность
избирательных механизмов антиоксидантной активности в условиях цепной реакции
свободных радикалов. Труды всеармейской научно-практической конференции
«Инновационная деятельность в вооруженных силах Российской Федерации».
МО РФ, ЛВО, Военная академия связи. Санкт-Петербург, 2009, с.332-334.
-
Родионов Г.Г., Плужников Н.Н., Листов
М.В., Мамыкин А.И. Экспериментальное моделирование патологических состояний
со свободнорадикальной этиологией и их математическое описание. Труды международной
конференции «Высокоинтенсивные физические факторы в биологии, медицине,
сельском хозяйстве и экологии». ФГУП Российский федеральный ядерный центр,
ВНИИЭФ, Саров, 2009, с. 88-97
-
G.N. Ling. A New Theoretical Foundation
for the Polarized-Oriented Multilayer Theory of Cell Water and for Inanimate
Systems Demonstrating Long-range Dynamic Structuring of Water Molecules
// Physiol. Chem. Phys. & Med. NMR, v.35, p. 91–130, 2003
-
Hubbard J. Electron correlations in narrow
energy bands. Proc. R. Soc. A, v. 281 p. 401, 1964
-
Anderson P.W.The resonating valence bond
state in La2CuO4 and superconductivity. Science, v. 235, p. 1196, 1987
-
Маркин В.С., Пастушенко В.Ф., Чизмаджев
Ю.А. Физика нервного импульса // УФН, т. 123, вып. 10, с. 289-332, 1977
-
Изюмов Ю.А. Магнетизм и сверхпроводимость
в сильно коррелированной системе // УФН т.161, вып. 11, с.1-46, 1991
-
Листов М.В., Мамыкин А.И., Кондаков А.Ю.
Селезнев А.Б. ИК-спектроскопия биологических жидкостей при моделировании
патологий со свободно-радикальной этиологией.
Комментарий
от авторов.
Эта статья была написана несколько лет тому назад, но мы не внесли в неё
никаких изменений. За истекшее с момента написания статьи время нам удалось
экспериментально подтвердить связь опистотонуса с концентрацией радикалов
в крови подопытных животных, регистрируя ИК спектры образцов крови, взятых
у мышей сразу после инъекции, и с некоторыми интервалами до одного часа
после инъекции. Действительно, экспериментальные точки ложатся на кривую,
описывающую изменение концентрации свободного радикала для конечного продукта
цепной реакции (см [2] в библиографии, а также Листов М.В., Мамыкин А.И.
Экспериментальное моделирование гемозависимого экзофтальма. Вестник Военно-медицинской
академии, вып.2 (42), 2013, с. 120-125). Читая статью, следует иметь в
виду, что модель Хаббарда в данном случае имеет отношение к формированию
нервного импульса в клетке, а рисунок 2
– к блокировке распространения
нервного импульса.
Комментарий редакции. Ортодоксальные сомнения
в применимости «твердотельного» математического
аппарата – всего лишь пережиток времён
«оДУХотворённой»
органической химии, когда считалось, что органика
– исключительно продукт деятельности живых организмов. Между тем, различие
органических веществ от минеральных, главным образом, заключено в том,
что первые синтезируются по «программе», а вторые
– по «образцу». Многие вирусы (тот же ВТМ
– вирус табачной мозаики) давно кристаллы, да и структурно
биомембраны, о которых идёт речь в статье, вполне подобны графану (легированному
графену), к которому использованный матаппарат безусловно применим, тем
более, что такое применение оказалось результативным.
 В оглавление
В оглавление