![]()
ПЕРИОДИЧЕСКАЯ СИСТЕМА ЖИВЫХ ОРГАНИЗМОВ
![]()
В последнее время человечество столкнулось с целым рядом необычных проблем. К ним относятся энергетические, информационные, экологические проблемы и ряд других. Появлению этих проблем предшествовала интенсивная деятельность человека в различных сферах своей жизни.
Очевидно, в связи с интенсификацией деятельности человека будут возникать и новые проблемы, прогнозирование которых представляет собой исключительно важную задачу. Однако, задача прогнозирования может найти свое решение только при изучении более широкого класса научных задач.
Одна из таких задач - задача о распределении энергии и ее взаимодействии с биологическими системами в космическом пространстве. В настоящее время, несмотря на то, что установлены многочисленные факты влияния космических сил (излучения Солнца, гравитационных возмущений от Луны и др.) на биологическую жизнь на Земле, космологические исследования не получили полноправного статуса научных исследований.
Несомненным является только то, что биологические процессы на Земле и космические процессы в Солнечной системе определенным образом связаны между собой. При этом нетрудно предположить и обратное влияние, то-есть влияние биологических процессов на Земле на космические процессы в Солнечной системе, которое может быть весьма значительным и существенно изменить наше понимание значения экологической деятельности человечества.
Одним из методов анализа указанной задачи является экстраполяция влияния на космические процессы дальнейшей эволюции живой природы и деятельности человека в рамках квазистационарной открытой термодинамической системы, которой является наша Земля.
Земля, как небесное тело с аккумулирующей энергию Солнца атмосферой, является системой, энтропия которой, на первый взгляд, должна неизбежно расти. Однако этого не происходит. Попробуем разобраться в возможных причинах этого явления.
С одной стороны, этому процессу препятствует излучение Земли и наличие эндотермических реакций н коре, а с другой - развитие живой природы, которая также потребляет и связывает энергию. Таким образом, полагая радиационное излучение собственно коры Земли величиной, при заданной температуре постоянной, можно выделить основные компоненты теплового баланса.
К ним относятся: вода в виде облаков, океанов и снега; эндотермические реакции в виде образования солей, минералов, нефти, угля и пр. и живая природа. Указанные составляющие тесно связаны между собой и образуют некоторую замкнутую систему, стабилизирующую поверхностный энергетический потенциал Земли.
Динамика саморегуляции приведенной схемы обеспечивается различными механизмами. Здесь направления потоков энергии совпадают с направлениями замкнутых колец и могут быть проиллюстрированы круговоротом воды в природе, действием живой природы на неживую природу, конвекционными потоками магмы и пр.
Механизм действия вулканов на энергетическую регуляцию очень убедительно изложен в гипотезе доктора Майкла Рампино из института космических исследований им. Годарда (США). В частности, как сообщает американский журнал "Сайенс дайджест", в случае охлаждения в короткий срок может увеличиться скорость пассатов, которые дуют в направлении, обратном направлению вращения Земли.
Если ветры начнут действовать в качестве тормоза, уменьшая скорость вращения нашей планеты, это приведет к тому, что магма под земной корой начнет двигаться в вихревых конвекционных потоках, что будет побуждать к извержениям вулканов. При этом изменится гравитационное взаимодействие Земли с другими планетами.Дальнейшее охлаждение приведет н подавлению вулканической деятельности, так как под давлением растущих ледников континенты начнут оседать. В то же время вода, связанная в виде льда, на континенте уменьшит давление на морское дно и оно начнет подниматься. Эти движения создадут участки напряженности в земной коре и приведут в движение магму, что усилит вулканическую деятельность. Тепло, выделяемое вулканами, вызовет активное таяние льда и испарение воды, что в виде облачности увеличит отраженный поток электромагнитной энергии и снизит конечный поток энергии, падающий на Землю, и, таким образом, приведет к дальнейшему похолоданию.
Влияние биологических процессов в данной схеме также можно проследить. Развитие растений на большой площади Земли влияет на динамическую составляющую ветров и пассатов, течений и волнообразований в Мировом океане. Это, в свою очередь, также влияет на скорость вращения Земли, что сказывается на гравитационном взаимодействии с планетами, спутниками и кометами. Таким образом, развитие растений и микроорганизмов существенно влияет как на вулканическую деятельность Земли, так и на коэффициент отражения Солнечного излучения, и, тем самым, на электромагнитную обстановку в Солнечной системе.
Особенно сложным в данном случае представляется процесс влияния животного мира, так как его изменение в этих случаях определенным образом отстает во времени от развития растительности, что, в свою очередь, формирует широкий частотный спектр различных (в том числе и космических) взаимодействий.
Анализ этих процессов невозможен без достаточно полного понимания структуры живых организмов как единой системы. Из литературы известно много попыток создания макросистемы живых организмов, однако, используемые авторами различные принципы расположения высших таксонов (типов животных, классов растений и пр.) не позволяли сформировать систему, подобную столбцам и строкам таблицы Менделеева. Это лишало полученные таблицы цельности и наглядности.
В настоящее время в связи с развитием биотехнологий, необходимости более глубокого изучения человека, как биологического вида, и обостренной экологической ситуации на Земле, значительные успехи в биологии возможны только при наличии единого взгляда на живой мир с точки зрения космизма биологии, его существенной связи с деятельностью человека. Естественно, с этой точки зрения уже нельзя удовлетвориться имеющимися макросистемами, в которых высшие таксоны не связаны в единое целое и не позволяют выявить основные корреляционные зависимости структурных, функциональных и энергетических признаков, аналогично периодической системе элементов. Тем не менее, во многих публикациях указывалось, что такая макросистема может быть создана.
Представляется, что основной причиной неудач предпринятых попыток является использование в качестве доминанты морфологических признаков строения живых организмов и их связей между собой без учета характеристик их взаимодействия и морфофизиологической дифференцировки [2].
В систематике традиционно используют филогенетический подход, основанный на приспособительном характере таксонометрических различий. Однако, в ряде случаев, наиболее существенные признаки не могут быть отнесены к признакам, унаследованным от общего предка, и не имеют видимого адаптивного значения.
Кроме того, аналогичные признаки возникают у таксонов, относящихся к различным уровням развития. Так, например, крылья у насекомых и птиц. Очевидно, что экофизиологический подход при описании "высших таксонов, как адаптивных типов" отражает экологическую структуру биосферы, но не учитывает общие законы полиморфизации, комбинирования и несовместимости.
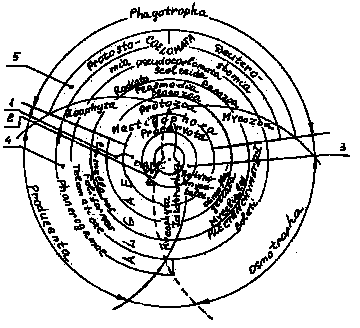
Рис. 1. Комбинированная четырехцарственная макросистема. Одноклеточные - 1; равноклеточные - 2; разно-клеточные, то-есть тканевые - 3; разнотканевые, то-есть тканеорганные - 4; централизованные - 5.
Попытки описать макросистему на экологической основе с использованием симметрий] велись издавна и последняя [2] также не дала необходимой наглядности, так как не имела физиологических признаков. С другой стороны, были попытки создания макросистем на основе учета метаболических и кибернетических систем по функциям организма и обслуживающих их органов [З], (см.рис.1). Последний подход представляется интересным с точки зрения использования модели Дж. Форрестола для описания живого организма.
Однако, структурно-функциональный анализ таких моделей был проведен только в кибернетическом аспекте, что не позволило автору выявить основные закономерности и построить непротиворечивую систему. Сущность используемого в данной работе подхода заключается во введении термодинамической модели живого организма в виде замкнутой фильтрующей оболочки с ядром, которую пронизывают потоки вещества и энергии, аналогично модели Дж. Форрестола для анализа взаимодействия организмов между собой и окружающей средой на основе структурно-энергетических критериев.
Взаимодействие окружающей среды с организмом в предложенной модели обуславливает сложноподчиненную иерархическую структуру строения живых организмов, изменяющуюся, как будет показано далее, периодически. Указанная особенность позволяет распределить высшие таксоны по строкам и столбцам, аналогично таблице Менделеева.
Рассмотрим возможные характеристики живых организмов, обладающие инвариантными относительно видов свойствами, которые могут быть положены в основу построения осей абсцисс и ординат в таблице. Робертом Уитткером предложена схема условного разделения живых организмов на три царства:
При этом данная схема основывалась на различных типах питания. У растений и грибов питание производится в основном за счет осмосорбционного транспорта растворов из окружающей среды при различных источниках энергии, а у животных - за счет заглатывания пищи.
В то же время указанные потоки питания не анализировались с точки зрения энергетики, а основное внимание уделялось наличию круговоротов веществ, то есть экологическому анализу. Тем не менее, анализ энергетических затрат на поддержание жизнедеятельности организмов за счет добывания пищи позволяет с другой точки зрения взглянуть на формирование макросистемы. Для наиболее общего анализа процессов взаимодействия организма с окружающей средой рассмотрим основные потоки вещества и энергии, пронизывающие организм.
Известно, что живые организмы синэргетически могут быть отнесены н квазистационарным открытым линейным термодинамическим системам, характеризующимся наличием векторов вещества и энергии, пронизывающих в разных направлениях данную систему. Известно также, что живой организм - это устойчивая саморегулирующаяся открытая система, обладающая высокой степенью организации.
Для обеспечения локального термодинамического равновесия в организме имеется система метаболизма и развитая гомеостазная система с наличием положительных и отрицательных обратных связей. При этом указанные потоки вещества и энергии живого организма можно условно представить в виде совокупности трех независимых векторов "А", "В", "С".
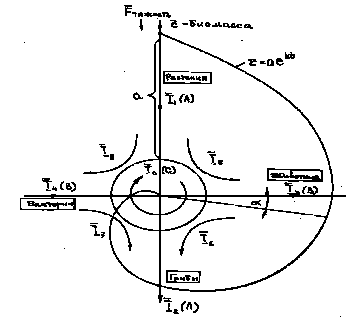
Рис.2 Потоки энергии и вещества.
Положим, что вектор "А" представляет собой поток вещества и энергии, пронизывающий живой организм (см. рис. 2). Через вектор "В" условно обозначим всю совокупность воздействий окружающей среды, а через ротационный вектор "С" - потоки вещества и энергии внутри организма, реализующие процессы обмена и обратных связей.
Для живых организмов вектор "А" представляет собой биофизический источник энергии и веществ компартмента, имеющего в своем составе: поток органической и неорганической пищи, воздуха, воды, электромагнитной энергии /солнечной, инфракрасной/или химической, а также поток выносимых из организма веществ и энергии.
Полагая квазистационарность рассматриваемой системы, вектор воздействия окружающей среды "В" отражает влияние на организм сил тяготения, климатических условий и других организмов и имеет свой адекватный противоположнонаправленный вектор "физиологический сток".
Ротационный вектор "С" в основном связан с процессами обмена и представляет собой потоки вещества и энергии внутри организма, условно он может быть назван вектором активации. В отличии от внешних векторов "А" и "В", вектор "С" для животных является внутренним и его содержание непосредственно связано со структурой организма.
Очевидно, что существование вектора активации "С" в кваэистационарной системе возможно только при наличии предохранительной оболочки, осуществляющей фильтрующую функцию. Кроме того, следует отметить, что квазистационарность вектора "С" в условиях внешних (например, климатических и др.) и внутренних (например, биологических и др.) воздействий обеспечивается наличием системы гомеостазных потоков и реакций. Таким образом, можно заметить наличие адекватной реакции на изменение каждого из указанных векторов.
У растений рассеянная электромагнитная энергия концентрируется в потенциальную химическую энергию, а у животных - концентрированная химическая энергия, поступающая в виде пищи, при движении рассеивается в окружающее пространство в виде электромагнитной энергии тепла. В свою очередь, внешнее воздействие гравитационных сил у растений компенсируется собственной накопленной в клетках потенциальной энергией, а у животных компенсируется изменением кинетической энергии при перемещении.
Рассмотрим взаимное расположение векторов "А" и "В" для различных организмов. Положим, что вектор "В" отображает действие силы тяжести. Тогда можно заметить, что для большинства растений вектора "А" и "В" лежат на одной прямой и совпадают по направлению, так как вектор "А" в основном представляет собой поток электромагнитной энергии. При этом угол a между векторами "А" и "В" равен нулю.
Для животных процесс добывания пищи связан в большинстве случаев с перемещениями вдоль поверхности земли и угол a тогда будет условно равен П/2.
Для грибов, питание которых производится за счет осмосорбционной транспортировки растворов из среды против сил тяготения, угол a условно равен П.
Отмирающие части растений и других организмов под воздействием силы тяжести падают на почву, содержащую гнилостные бактерии, и служат для последних пищей. Колонии бактерий растут в основном горизонтально, что позволяет угол a условно положить равным 3/2П или - П/2. Таким образом, при плавном вращении векторов "А" и "В" можно рассмотреть все известные царства организмов, включая переходные формы.
Если положить среднее количество ежегодно обновляемой биомассы всех организмов условно в виде длины вектора "А", то распределение биомассы организмов между царствами можно представить в виде спирали, шаг которой равен переменной части биомассы растений; площадь под спиралью в первом квадранте, (см. рис.2) будет равна площади под спиралью во втором и третьем квадранте (с точностью примерно 9%), то-есть интегральное значение биомассы растений будет равно интегральному значению переменной биомассы остальных организмов.
Биомасса организмов (примерно 9%), реализующих фотосинтез, но не являющихся растениями, расположена в четвертом квадранте, что также соответствует схеме на рис.2. Указанными организмами в основном являются одноклеточные (прокариоты и эвкариоты).
Принятая схема достаточно наглядно иллюстрирует соответствие между всеми известными царствами в растительном и животном мире, то-есть царствами растений, животных, грибов и бактерий. Заметим, что величина вектора активации "С" меняется взаимооднозначно в соответствии с изменением уровня сложности организма и носит дискретный характер. В соответствии с законом сохранения энергии, все функции живого организма, требующие затрат энергии, должны в конечном счете осуществляться за счет внешних источников энергии, из которых основным является солнечная энергия, служащая первичным источником для автотрофных растений, стоящих в начале пищевых цепей.
Из общей энергии солнечного излучения Земли достигает лишь ничтожная доля (радиус Земли составляет чуть больше 4-х стотысячных радиуса ее орбиты - телесный угол оцените сами). Из этого количества энергии 30% отражается поверхностью атмосферы, а по пути через атмосферу поглощается еще 25%, так что поверхности Земли достигает всего лишь около 45% доли солнечного излучения, излучаемой Солнцем в этот ничтожный телесный угол.
Общая энергия излучения, падающего на поверхность Земли, примерно равна 80000 ТДж/с (Тераджоулей в секунду). Около 45% этой энергии приходится на спектральную область между 380 и 740 нм, и только эта часть энергии может использоваться растениями для фотосинтеза. Таким образом, энергия фотосинтетического активного излучения составляет ~ 36000 ТДж/с.
Минимальная величина ротационного вектора "С" для растений определяется с одной стороны "красной" границей электромагнитного излучения, а с другой - энергией молекул АТФ (аденозинтрифосфата) (Wатф = 30 кДж). Энергия активации молекулы в соответствии с формулой Энштейна находится из выражения Wa = hv, где
Отсюда можно определить максимальный темп синтеза молекул АТФ, являющихся основным энергетическим носителем в биологических организмах. Он не превышает 0,1 сек. Например, у человека каждая молекула адеэинтрифосфата (АТФ) расщепляется и вновь регенерируется 2400 раз в сутки, так что средняя продолжительность ее жизни менее 1 минуты, у других живых организмов есть и еще меньше.
Это подтверждает правомерность использованного подхода для анализа энергетических характеристик живых организмов. Учитывая различный уровень организации живых организмов, энергетическое соотношение между векторами "А", "В" и "С" можно выразить в виде A + B = kС, где k -характерный коэффициент энергетики организма данного вида; C = nФ, где Ф = соnst - число АТФ (примерно 30 кДж/моль), n - комплексный коэффициент.
Как было сказано ранее, для устойчивого существования вектора активации "С" и стабилизации вектора "А" рассматриваемая система должна содержать замкнутую фильтрующую оболочку. Указанная оболочка защищает организм и обеспечивает его жизнедеятельность путем регулирования потоков вещества и энергии, пронизывающих организм.
Фильтрующая функция оболочки образуется в основном за счет радиальной анизотропии пропускающих свойств ее стенок, которую можно условно оценить различной степенью упорядоченности ее элементов, то есть энтропией. Отсюда в большинстве случаев энтропия внутренних и внешних поверхностей фильтра различна.
Например, широко известно явление избирательной транспортировки молекул веществ оболочками (мембранами) клеток за счет различной геометрической формы внешней и внутренней поверхности стенок. Учитывая квазистационарность процессов в рассматриваемых открытых системах, можно заметить чрезвычайно важное свойство, заключающееся в том, что указанные оболочки с точки зрения биологического взаимодействия являются замкнутыми и не пересекаются между собой.
Таким образом, модель открытой системы представляет собой условно замкнутую оболочку, содержащую стенки, обладающие свойством радиальной анизотропии, а в ее полости имеется ядро, которое ответственно за преобразование энергии и вещества.
Учитывая энергетическую квазиуравновешенность организма, естественно предположить зависимость структуры ядра от строения оболочки. Тогда, полагая, что множество Sо отражает энтропию строения поверхности оболочки, а множество Sя -энтропию ядра (как меру разнообразия структуры элементов ядра), необходимо ввести дополнительное множество Sд , отражающее взаимодействия организма с внешней средой.
При этом можно отметить два случая. В первом - сложное устройство ядра определяет сложную структуру внутренней поверхности оболочки и относительно разнородную - наружной поверхности оболочки. В этом случае можно говорить о сравнительно небольшом возможном разнообразии пищи (вектора "А") и достаточном разнообразии допустимых воздействий окружающей среды (вектор "В").
Во втором случае наблюдается обратная картина (см. рис.3). Таким образом, введенное ранее дополнительное множество Sд в определенной степени характеризует разность энтропии векторов "А" и "В".
Например, у прокариот (безъядерных одноклеточных организмов) наблюдается довольно сложное устройство оболочки с относительно однородным содержанием ядра в виде протоплазмы, что позволяет им относительно эвкариот (одноклеточных организмов с ядром) иметь большее разнообразие в пище, но меньшее в среде обитания. Из этого можно сделать вывод, что у эвкариот устройство организма дуально противоположно прокариотам с точки зрения введенных энтропийных оценок.
Для более сложных организмов внешней средой для каждого уровня иерархии его органов является последующий иерархический уровень организации организма, который, соответственно, отличается от предыдущего этропийно на величину Sд.
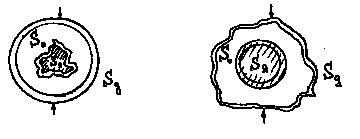
Рис. 3. Зависимость структуры ядра от строения оболочки
Отсюда, учитывая ранее описанное свойство непересекаемости и наличие группы радиальной изотропии оболочек, следует, что внутреннее строение всех живых организмов характеризуется периодичностью функции изменения разнообразия устройства внутренних органов (энтропией) и может быть описано топологической структурой их связей в виде "дерева".
Например, у млекопитающих относительно однообразные по устройству клетки образуют в достаточной степени разнообразные органы (сердце, печень и др.), объединенные относительно однообразной внешней оболочкой (шкурой). Для дальнейших рассуждений можно ввести некоторую условную классификацию элементов обобщенной модели организма:
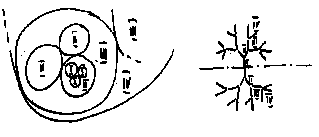
Рис.5. Планетарная структура "дерева". Рис.6 Радиальная структура "дерева".
Большинство живых организмов могут быть представлены в виде топологических моделей типа "дерева" с последовательным и параллельным соединением ветвей (рис.5, рис.6), для которых законы сохранения могут быть описаны следующими уравнениями:
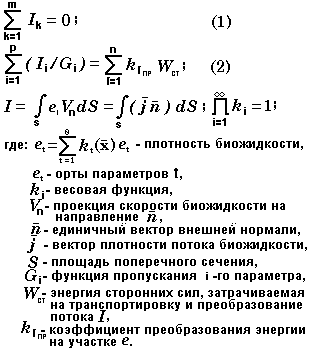
При этом использовались следующие понятия и определения, введенные автором:
Отметим некоторые свойства рассматриваемой обобщенной модели организма:
Для оболочек наблюдается обратная картина. В ходе развития организма растет многообразие комбинаций функций, которые интегрируются и ведут к новым, обычно более сложным, системным уровням. Это повышение уровня касается организации и приводит к уменьшению энергетических, материальных и информационных затрат, к более рациональной стратегии размножения и сохранения вида, к созданию предпосылок увеличения ареала, существования.
На повышение организации может указать уменьшение числа однородных структур или дифференциация исходно одинаковых структур, ведущая к разделению функций. Таким образом, имеют место две основные тенденции эволюции. Рационализация структуры и функции, например, путем концентрирования, централизации или функционального объединения (синорганизации) каких-либо элементов и усложнение, связанное с дифференциацией и разделением функций.
Результатом усложнения является возрастание пластичности структуры и функций, способствующих уменьшению зависимости от флуктуаций параметров окружающей среды, Доминирующим параметром на Земле можно считать действие космичесних сил, таких, как гравитация и солнечное излучение, рассмотренные ранее в обобщенной модели живого организма.
Очевидно, что они взаимосвязанно с рассмотренными тенденциями интегрирования (объединения, суммирования) и дифференцирования (непересекаемость) органов и функций, которые являются необходимыми и достаточными условиями для образования систем, структура которых может быть описана в виде графа типа "дерево" радиального или планетарного класса.
Приведенные свойства модели позволяют предположить основное фундаментальное свойство, заключающееся в периодическом дискретном изменении свойств организмов в зависимости от изменения сложности их структуры. Отметим, что рассмотрение организма в отрыве от характеристик окружающей среды не позволяет производить достаточно полную оценку его основных свойств.
Анализируя поведение животных, можно заметить, что все они перемещаются, и, таким образом, обладают некоторой кинетической энергией. Полагая, что взрослая особь расходует свою энергию в основном на движение, можно оценить поток энергии, пронизывающий их организм в среднем за сутки в период зрелого возраста (когда прекратился процесс размножения, не строится жилище, не накапливаются жиры и т.п.).
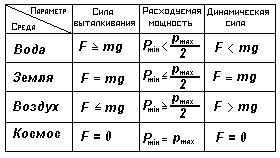
Допустим, что в этот период основные затраты энергии идут на добывание пищи, поддержание постоянной температуры тела и рассеяние тепла в окружающую среду. Заметим, что на всех животных действует сила тяжести, в основном определяющая сопротивление окружающей среды движению организма. Однако, находясь в разных фазовых средах, организм подвергается разному действию сил Архимеда, компенсирующих в определенной мере действие сил гравитации (см. табл.1).
Например, плотность организмов, находящихся в воде, приближается к плотности воды, за счет чего пелагические рыбы при перемещении испытывают в основном только сопротивление воды. У животных, плотность организмов которых несколько превышает плотность воды, движение в основном происходит вдоль и по поверхности дна. При этом для своего перемещения организм приподнимается над поверхностью дна частью своего тела (за счет ног или изгибания туловища) и переносит его отдельными частями в процессе движения.
Аналогичным образом происходит перемещение животных на поверхности суши. Это требует при перемещении по дну затрачивать энергию на перемещение в среднем несколько большую, чем при перемещении в пелагической зоне. Соответственно, средняя энергия перемещения земноводных существенно больше средней энергии перемещения донных животных, а у животных, перемещающихся над поверхностью земли, больше, чем у земноводных.
При перемещении животных в воздухе им приходится тратить энергию на поддержание своего организма в воздухе, почти полностью компенсируя действие гравитационных сил, так как величина архимедовой силы в воздухе весьма мала, вследствие большой разницы между плотностью тела организма и воздуха.
Анализ энергетических затрат опорно-двигательных функций живых организмов в поле тяготения при наличии различных фазовых сред показывает связь анатомического строения организмов с дифференцированием основных функций.
Таблица 1.
Проведенный анализ зависимости энергетических затрат (или потребления кислорода) при движении различных млекопитающих от их массы показал, что в логарифмических координатах эта зависимость представляет собой прямую линию [3]. Это говорит о постоянстве для данного таксона скорости изменения удельного потока энергии от массы видов:
dWn/dMn = - C, где:
Ввиду того, что на всех млекопитающих, двигающихся по земле, гравитационная сила действует одинаково, изменение потока энергии происходит только в зависимости от массы тела, то-есть в зависимости от силы гравитационного взаимодействия живого организма с землей.
Таким образом, анализ действия гравитационных сил показывает на различную величину усредненных энергетических затрат на перемещение организма в различных фазовых средах, что позволяет условно разделить их на некоторые группы по энергетическому признаку.
В силу того, что животные покрывают свои энергетические потребности, поедая растения или других животных, различная величина средних энергетических затрат определенным образом характеризует различные биологические взаимодействия между типами и царствами. Под биологическим взаимодействием в дальнейшем будем понимать среднюю величину энергетического потока, пронизывающего организм.
Очевидно, при такой формулировке абсолютная масса потока пищи у разных организмов может быть различной за счет разброса в калорийности и различного коэффициента ее усвоения. Кроме того, следует отметить, что влияние гравитационных сил на простейшие организмы несколько ослаблено за счет действия различных электрических и молекулярных сил. Этим можно объяснить значительную широту сфер обитания простейших организмов.
Таким образом, усредненный поток энергии, пронизывающий организм животного, может быть различным и в основном зависит от свойств организма и качества пищи.
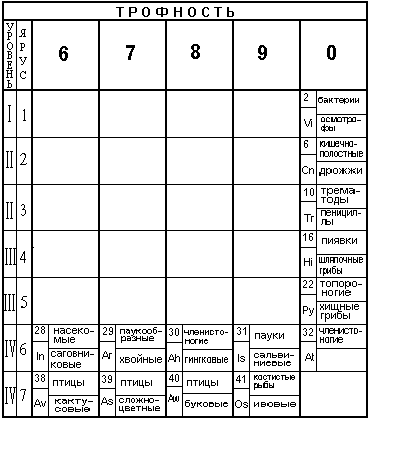
Назовем способность организма обеспечивать свою энергетику за счет пищи трофностью. По числу возможных фазовых сред обитания живых организмов введем шкалу трофности, содержащую девять единиц. За единицу трофности примем средний удельный поток энергии, проходящий через одну клетку одноклеточного организма, находящегося в воде.
Изменение трофности, очевидно, будет происходить при появлении в организме клеток с новым типом строения, которые образуются за счет дифференцирования функций отдельных органов. При этом функции организма, связанные с добычей пищи и теплорегулированием, являются доминирующими и их энергетика по отношению к полной энергетике организма достаточно полно характеризует свойство предпочтительной трофности таксона.
Например, паразиты не затрачивают энергию на добычу пищи и имеют с точки зрения введенного определения трофность, равную нулю. Рассмотрим разные уровни организации живых организмов, распределенных по уровням иерархии. Каждый уровень иерархии содержит представителей животных и растений, расположенных соответственно введенному понятию трофности.
I уровень иерархии содержит четыре класса простейших животных:
Следует заметить, что в классе жгутиковых имеются эвглены, в клетках которых имеются хлорофилловые элементы. Растения представлены одноклеточными водными организмами из отдела водорослей: синезеленые водоросли, золотистые водоросли, известковые водоросли, диатомо-вые водоросли, желтозеленые водоросли, криптофитовые и динофлагеляты. Там же представлены эвглены.
II уровень иерархии. В нем имеется три типа многоклеточных животных:
Отметим, что известковые губки отличаются от других классов наличием известковых спикул. Устройство всех трех типов содержит набор из соответственно трех, четырех и пяти видов строения клеток (стрекальные, нервные, пищеварительные, кожно-мускульные и яйцевые /сперматозоидные). Растения занимают три клетки трофности соответственно многоклеточными водорослями:
III уровень иерархии также содержит три типа животных:
плоские черви, включающие в себя три класса:
Последние два класса представляют собой паразитов:
В их устройстве появляются клетки с новым строением, из которых строится кожа, кишки и глаза. Растения содержат также три класса водорослей:
IV уровень иерархии организмов животных состоит из трех типов:
При этом тип кольчатых червей включает класс пиявок, класс малощетинковых кольчецов и класс многощетинковых кольчецов. В этом уровне иерархии животные имеют новые виды строения клеток (клетки параподий, хитиновые щетинки, сенсорные щетинки, кровеносные каналы и нервные узлы).
Растения из трех отделов располагаются соответственно:
V уровень иерархии представляет собой пять классов моллюсков:
У животных этих классов появляются клетки новых видов строения, из которых состоят: желудок, сердце, половые органы, зубы, легкие, печень и язык.
Растения представлены из отделов высших (сосудистых, листостебельных) растений, в частности, отделов папоротниковидных и плауновых. Здесь классы:
VI уровень иерархии содержит два типа животных:
имеющие новые органы из клеток новых видов строения: ядовитые железы, панцирь, органы слуха, крылья, волоски, паутинные железы, жабры, пищеварительные железы, половые железы, зеленые железы, легочные мешки, мальпигиевы сосуды и жировое тело.
Растения представлены из отделов папоротниковых и семенных растений. В том числе из классов:
VII уровень иерархии состоит из животных типа полухордовых и типа хордовых. Последний тип состоит из трех подтипов (личинкохордовые, ланцетники и позвоночные) и семи классов:
Указанные животные из клеток нового вида строения имеют следующие органы: эндокринные железы, селезенка, желчный пузырь, легкие, мозг, уши, органы обоняния и вкуса, щитовидная железа, волосы, перья, чешуя, молочные железы, гормональные железы, кости скелета, потовые железы и половые органы.
Растения представлены классами и подклассами отдела семенных растений. Соответственно, они расположены по шкале трофности в соответствии с подклассами: водокрасовые, рясковые, частуховые, рдестовые, наядовые; лилейные, орхидные, бромелиевые, ситниковые, злаки; бобовые, камнеломковые, розоцветные, зонтичные, молочайные; кактусовые, гвоздичные, мариевые, гречишные; сложноцветные, горечавковые, маслиновые, пасленовые, коричниковые, губоцветные; буковые, березовые, тутовые, коноплевые, крапивные; ивовые, фиалковые. Как видно, они представлены крестоцветными, мальвовыми классов двудольных и однодольных подотдела покрытосеменных.
Колонка трофности "0" содержит паразитов и грибов, представленных соответственно:
Таблица 2 (часть 1).
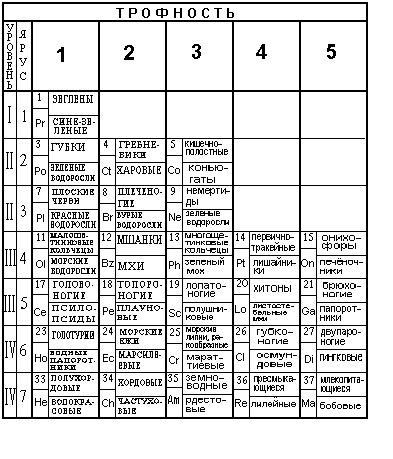
Таблица 2 (часть 2).
Рассмотрим проявление подмеченной закономерности для классификации объектов живой природы. Эволюционное изменение среды обитания живых организмов из воды на землю и воздух ограничено возможными для организмов данного уровня иерархии удельными энергетическими затратами клеток, увеличивающимися по мере усложнения их структуры.
Изменение cреды обитания организмов наглядно отражает общее изменение свойств организмов. Учитывая значительное число известных в настоящее время видов организмов, как и ранее ограничимся при рассмотрении наиболее крупным делением живого мира на типы, приводя по мере необходимости примеры отдельных видов.
Принцип построения периодической системы типов живых организмов заключается в выделении определенных периодов и групп (расположенных, соответственно, горизонтально и вертикально).
Положим, что период - это совокупность высших таксонов, содержащих виды, средой обитания которых является вода, а заканчивающихся - паразитами, средой обитания которых являются организмы с более высоким уровнем сложности.
Каждая группа подразделяется на две подгруппы, причем, организмы первой подгруппы принадлежат животному миру, а второй - растительному, и таксоны обеих групп имеют одинаковый иерархический уровень организации. Особое значение для понимания целостности мира живого имеет понимание глубокого дуализма между царствами растений и животных, что позволяет объединить оба эти царства в рамках одной макросистемы.
Анализ его показывает, что разные периоды соответствуют наличию у организмов различных ярусов гомеостаза, что объясняет ослабление значения естественного отбора с увеличением сложности вида и, соответственно, увеличение устойчивости вида.
С увеличением номера таксона также растет информационная сторона жизни относящихся к нему организмов. Таксоны, помещаемые в таблицу, информационно изолированы между собой. Для животных периодический закон изменения числа сфер обитания R от степени сложности животных может быть описан уравнением
![]() где
где
Рассмотрим предложенную возможность систематизации животных по признакам трофности и уровням организации. Под уровнем организации будем понимать уровень иерархии организма, определяемый числом независимых роторов векторов активации в рассмотренной ранее планетарной системе (ядро и оболочка).
Очевидно, первому уровню иерархии соответствуют простейшие одноклеточные животные - эвкариоты, у которых величина трофности, в соответствии с введенным определением, равна единице. Прокариоты не относятся к рассматриваемому царству и могут быть классифицированы самостоятельно.
Заметим, что второй и третий уровень иерархии содержит животных с трофностью от 1 до З, а четвертый и пятый уровни - с трофностью от 1 до 5.
Аналогично, шестой и седьмой уровни иерархии содержат животных с трофностью от 1 до 9.
Нулевому значению трофности соответствуют паразиты. Легко заметить, что величина трофности определяет среду обитания и определенным образом отражает кинетические характеристики животных.
Увеличение уровня иерархии связано с увеличением сложности организмов, входящих в соответствующие периоды, и имеющих сходное анатомическое строение. Например, если животным, относящимся к первому уровню иерархии, в большинстве случаев соответствует деление, как основной способ размножения, то второму уровню - почкование, третьему, четвертому и частично пятому - гермофродитное, а пятому, шестому и седьмому - половое.
Аналогично, у животных в первом периоде пищеварение обеспечивается пищеварительной вакуолью, во втором периоде появляется кишечная полость, в третьем - кишки, в четвертом - пищевод, в пятом - желудок и язык, в шестом - органы внешнего пищеварения, в седьмом - система органов пищеварения и желчный пузырь.
Нервная деятельность рассмотренных групп животных на первом периоде ограничивается раздражимостью, на втором периоде переносится на нервные клетки; на третьем периоде появляются нервные стволы; на четвертом периоде появляется нервный цикл; на пятом уровне - нервные узлы; на шестом уровне образуется нервная система, а на седьмом уровне появляется центральная нервная система, включающая мозг.
Оболочка рассмотренных организмов эволюционирует от оболочки клетки соответственно к клеточной оболочке и далее, через образование отдельных мышц к системе мышц; образование защитной раковины, панциря с волосками, и на седьмом периоде образуется чешуя, волосы и перья.
Аналогичным образом можно показать усложнение сенсорных систем, систем защиты, крови, лимфы, воздухообмена, органов перемещения и др. Все выделенные типы и классы животных имеют порядковый номер, определенным образом соответствующий числу принципиально различных типов строения клеток, из которых состоят их организмы.
Величина трофности для животных каждого выделенного таксона может быть различной, но в таблице даны преимущественные значения трофности, характеризующие таксон в целом. На животных, относящихся к таксонам, расположенным в начале таблицы, можно заметить, что доминирующее влияние на устойчивость оказывает естественный отбор. Указанная доминанта постепенно переходит к концу таблицы в развитый гомеостаз, обеспечивающий устойчивость сложного организма.
Кроме того, выделенные таксоны можно охарактеризовать информационной изолированностью между собой и энергетическим сродством близлежащих таксонов. Анализируя таблицу, легко заметить периодическое изменение среды обитания по мере их усложнения, что ставит под сомнение гипотезу о выходе живых организмов из океана на поверхность суши в процессе эволюции.
Это сомнение усиливается монотонностью функции, описывающей зависимость времени эволюции типа организма от номера таблицы. Изменение среды обитания таксонов по мере усложнения организмов чередуется с изменением ареала их обитания, что обусловлено приспособленностью оболочки животного к условиям внешней среды.
Особое положение занимает пятый период, в котором животные создают себе внешнюю оболочку из омертвевших клеток собственного организма. Впоследствии для образования внешней оболочки используются омертвевшие части других организмов /гнездо/ и предметы неживой природы.
Кроме того, как уже отмечалось, число различных гомеостазных реакций в организме определенным образом связано с номером периода. При этом характерна закономерность изменения максимального числа трофности на нечетных периодах, что объясняется свойствами четности изотропии внешних оболочек.
В силу неизменности потока солнечной энергии, пронизывающего Землю, каждый уровень иерархии постоянно заселен, строго определен и замкнут. Таким образом, на базе сформированной ранее обобщенной синергетической модели живого организма удалось выявить аналогичную Менделеевской периодическую структуру макросистемы животных и растений.
Периодический закон заключается в периодическом изменении среды обитания животных по мере изменения сложности организма, определяемой функцией изоморфизма клеток в совокупности с изменением числа ярусов гомеостаза и последовательным переходом последнего от преимущественно параметрического к функциональному.
Полученная закономерность может иметь большое методологическое значение при медико-биологических исследованиях, в учебном процессе и понимании экологических проблем в космическом масштабе.
Рассмотренная обобщенная модель была бы неполной без анализа особенностей макроклассификации растений. Структура "дерева", наблюдающаяся у растений, реализуется с помощью "узлов" расходящихся потоков биожидкости, то есть интегральной функции, аналогичной объединению общей оболочкой.
При этом угол между векторами потоков характеризует отношение векторов "А" и "С", являющееся индивидуальной особенностью таксона. В случае, когда разветвление сопровождается дифференцированием основной функции органа, происходит изменение уровня иерархии всего таксона. Таким образом, в общем случае, имеет место периодическое изменение удельных энергетических характеристик /кинетических для животных и потенциальных для растений/ по мере изменения структуры сложности организма.
Более строго эту закономерность можно сформулировать следующим образом: закономерность периодического изменения удельных энергетических характеристик в онтогенезе и филогенезе живых организмов заключается в том, что эволюционное изменение области обитания происходит периодически в зависимости от уровня морфологической интеграции в совокупности с функциональной дифференцировкой.
Заметим, что изменение уровня организации взаимооднозначно связано с явлением редукции разнообразия видов, характеризуемой уменьшением числа форм по мере усложнения организмов. В итоге можно отметить, что выявленные закономерности рассмотренной модели могут явиться основой для самостоятельного изучения в рамках конструктивного направления в биологии.
Но помимо этого, аналогичная периодическая система может быть построена для технических объектов, где аналогом растений являются здания, а аналогом животных - транспортные технические средства. При этом ряд транспортных средств с самостоятельной системой жизнеобеспечения может быть выделен в качестве самостоятельных таксонов.
Исследование данной проблемы позволило создать обобщенную синергетическую модель живого организма, сформулировать основные закономерности ее развития и выявить необходимость условия формирования макросистемы живых организмов, которая имеет аналогичную Менделеевской периодическую структуру. В ней выявились некоторые основные связи между характеристиками биосферы, строением организмов и структурой ДНК.
Анализ полученной макросистемы показывает непротиворечивость известным в настоящее время закономерностей и позволил заметить новые, связанные с экофизиологическими свойствами живых организмов. Полученная макросистема может служить основой для локального и глобального анализа экологических систем и возможных в них отклонений, например, в процессе экодиагностики [1]. Например, если наблюдать одновременно мощность излучения определенной Звезды и отраженную от поверхности планеты этой Звезды мощность ее излучения, то при наличии биологической жизни на этой планете можно с достаточно высокой степенью достоверности утверждать, что эти параметры будут вести себя как комплексные величины.
Допустим, что мощность излучения звезды изменилась во времени (мощный протуберанец) за период, сопоставимый с периодом обращения этой планеты вокруг Звезды. Тогда поглощение рассматриваемой планетой энергии Звезды будет изменяться с запаздыванием и иметь сложный волновой характер. Таким образом, альбедо планеты будет иметь спектр собственных частот.
Как выявлено ранее, существование биологических организмов на Земле определенным образом связано с кинетическим параметрами движения Земли, и, таким образом, с гравитационными процессами в Солнечной системе. При этом энтропийная направленность деятельности живых организмов в определенной мере обеспечивает космический "гомеостаз" (устойчивость) планет и других небесных тел Солнечной системы, а также электромагнитную обстановку в нашей Галактике.
Вскрытые при анализе разработанной макросистемы закономерности позволяют перейти к детальному анализу значения биологических систем в космических взаимодействиях. Особенно важно отметить, что воздействие Человека на экологическую обстановку на Земле в настоящее время не учитывает необратимые процессы, проходящие в космосе, и, тем самым, недооценивают последствий от экокатастроф на Земле.
Значение полученных закономерностей в том, что они позволяют анализировать глобальные события не только происходящие на Земле, но и в ближнем и даже дальнем космосе. Это дает возможность уже сегодня предложить несколько (оптических, гравитационных, электромагнитных) методов обнаружения существования биологических объектов в других звездных системах и оценить их уровень эволюции.
Кроме того, весьма актуальными являются астрономические наблюдения, которые могут экспериментально подтвердить многие теоретические положения предлагаемой космогонической теории.
Заключение. В результате анализа разработанной обобщенной термодинамической модели живых организмов осуществлена первая попытка формирования макросистемы живых организмов на основе структурно-функциональных признаков. Достоверность предложенной классификации обеспечивается в определенной мере непротиворечивостью и монотонностью в ней известных закономерностей. Наглядность и целостность макросистемы, несмотря на сложность отдельных ее положений, позволяет предположить ее значительную методическую и научную ценность. Дальнейшее уточнение введенных определений и таксонов представляет собой самостоятельную научную задачу систематики, решение которой позволит перейти к анализу тонкой структуры системы и ее эволюции.
ЛИТЕРАТУРА
Куэьмин П.П. Периодическая система видов живой природы..В сб. "Метрологическое обеспечение и стандартизация".Тезисы докладов VII Всесоюзной школы по проблемам метрологического обеспечения и стандартизации. Фрунзе, 1989.
Чайковский Б.В. Опыт экофизиологической макросистемы. Сб.статей "Методы исследования в экологии и этологии" института эволюционной морфологии и экологии животных им. А.Н.Северцева АН СССР, Пущино, 1986.
Новосельцев В.Н. "Организм в мире техники: кибернетический аспект", М., Наука, 1989.